

 蛋白酶-切除蛋白标签的有力工具
蛋白酶-切除蛋白标签的有力工具
14857
2023-10-17
随着蛋白质组学技术的迅猛发展,重组蛋白的使用在近年来大大增加,为了提高重组蛋白的产量或赋予重组蛋白新的特性(这些特性可以用来对重组蛋白进行纯化或检测),可以在克隆过程中添加额外的标签到目标蛋白的N端或者C端,标签可根据功能分为两类,纯化标签和可溶性标签,纯化标签可用于亲和层析,快速纯化蛋白;可溶性标签用于促进蛋白质的正确折叠,增加可溶性。
常见的标签有6*His、GST、MBP、Strep(Ⅱ)、Flag、SUMO和组合式双标签或多标签。GST、MBP、SUMO等蛋白标签由于分子量较大,通常需要去除,以消除其对目标蛋白的结构与功能的影响;多肽标签(如Strep(Ⅱ)、Flag)分子量较小,免疫原性较低,基本不影响蛋白质的结构和功能,通常可以不用去除,但用于结构生物学或其他较高级研究的蛋白通常也要将小分子量多肽标签去除,以达到最佳效果。实际操作中化学切割方法较为少用,因为不仅裂解位点的特异性低,还可能对目的蛋白产生不必要的修饰。目前常用的是反应条件温和且具有高度的位点特异性的酶解法。
表1. 常用的蛋白酶及对应切割位点
|
蛋白酶 |
表达宿主 |
切割位点 |
酶的作用 |
|
毕赤酵母 |
Asp-Asp-Asp-Asp-Lys |
去除位于蛋白N-末端的融合蛋白,以除去不需要的标签 |
|
|
大肠杆菌 |
Glu-Asn-Leu-Tyr-Phe-Gln↓-Gly |
融合蛋白标签切除 |
|
|
大肠杆菌 |
Leu-Glu-Val-Leu-Phe-Gln↓Gly-Pro |
特异切割含有3C酶切位点的融合蛋白 |
|
|
大肠杆菌 |
识别完整的含有100个氨基酸的SUMO标签蛋白 |
高效地把SUMO从融合蛋白上切割下来 |
|
|
牛的血浆纯化制得 |
Leu-Val-Pro-Arg-↓-Gly-Ser和Gly-Arg-↓-Gly |
融合蛋白标签切除 |
克隆过程中构建相应的氨基酸识别序列,即可保证后续可通过合适的蛋白酶对融合蛋白标签进行切除。
小翌已为大家准备好了性价比极高的几款蛋白酶,下面带大家一一了解下:
重组肠激酶(rEK)是一种高纯度的重组牛肠激酶轻链片段,其氨基酸序列与牛源肠激酶轻链一致,有着和天然提取的肠激酶同样特异的切割酶活性,切割位点是Asp-Asp-Asp-Asp-Lys(DDDDK),可用于去除位于蛋白N-末端的融合蛋白,以除去不需要的融合标签,保障重组融合蛋白准确的N端序列,同时重组肠激酶具有比天然酶更高的切割活性,是N末端融合通常选择的蛋白酶,在其他残基的不规则切割发生水平较低。
翌圣重组肠激酶(His标签)为采用毕赤酵母分泌表达的高纯度、高活性、高特异的牛肠激酶,无外源性的病毒污染,生产过程不使用任何动物源原料,可以在较宽pH范围(4.5-9.5)和较宽温度范围内有效切割融合蛋白且在各种去垢剂和变性剂存在的条件下仍具有部分活性。本品带His标签,在切割反应完成后,可通过Ni2+亲和柱轻松去除,极大地简化了后续的纯化流程。
凝血酶是由大小分别为31 KD和6 KD的两条肽链通过二硫键组成的一种丝氨酸蛋白水解酶,可从动物血浆中利用已激活凝血酶原水解制得;凝血酶切割序列专一,水解效率高,在分子生物学、生物化学或生物工程制药研究中常作为工具酶用于重组融合蛋白(包含凝血酶识别位点)的特异性断裂。
翌圣牛凝血酶是从牛的血浆纯化制得,具有纯度高、比活高、不含有其他蛋白酶活性、灭活病毒、生产稳定性好等特点。该酶最适切割位点:A-B-Pro-Arg-↓-X-Y (其中,A和B为疏水氨基酸,X和Y为非酸性氨基酸),常见的识别序列为:1. Leu-Val-Pro-Arg-↓-Gly-Ser。2. Gly-Arg-↓-Gly。凝血酶可从切割产物中用苯甲脒或肝素琼脂糖亲和纯化去除。
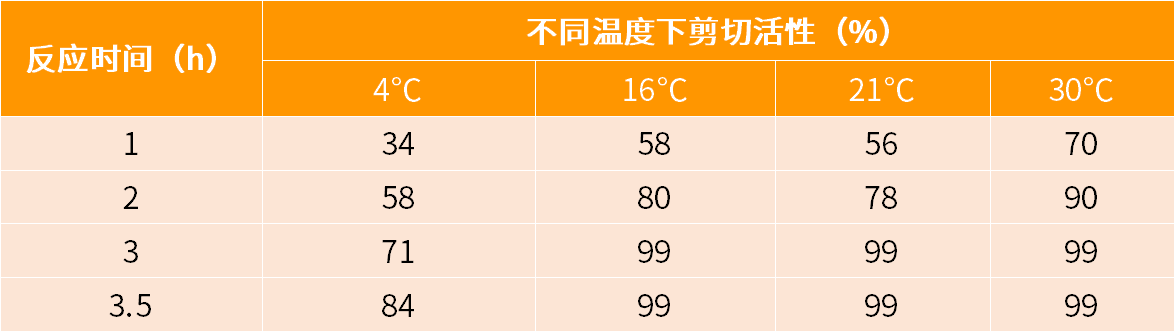
重组型TEV蛋白酶(rTEV)是经过基因工程改造和纯化后的重组蛋白酶,不仅保持天然TEV酶的功能活性,且在广谱的温度范围内表现出更强的稳定性和特异性。rTEV是一种用来切除融合蛋白上亲和标签的常用工具酶,具有很强的位点特异性,严格识别七氨基酸序列EXXYXQ↓(G/S),切割位点在谷氨酰胺和甘氨酸/丝氨酸之间。普遍应用的七氨基酸序列为:Glu-Asn-Leu-Tyr-Phe-Gln↓-Gly。rTEV在pH 7.0,30℃时可达到最佳活性,但在pH 6.0-8.5和温度4-30℃的广泛范围内皆有活性(见表2),使得反应条件的选择可根据目的蛋白的情况而灵活变动。另外rTEV切割后也能利用其N端的6×His标签,通过Ni-NTA树脂去除,以达到纯化目的蛋白的目的。
表2. 不同温度下rTEV酶切活性
3C蛋白酶是切割小RNA病毒科非结构蛋白的关键酶,在病毒复制过程中发挥着重要作用。鼻病毒属于小RNA病毒科,其中的人鼻病毒3C蛋白酶基因编码区全长552bp,编码的蛋白质相对分子质量约为22000 Da。人鼻病毒3C蛋白酶具有高度的酶切特异性,能特异切割位于Gln-Gly之间的肽键,识别位点为 Leu-Glu-Val-Leu-Phe-Gln↓Gly-Pro。
翌圣将人鼻病毒3C蛋白酶的编码区基因,在大肠杆菌中进行重组表达,纯化后获得了高纯度的重组3C蛋白酶,该蛋白酶能特异切割含有3C酶切位点的融合蛋白,具有良好的生物学活性。同时,翌圣生产的3C蛋白酶带有GST和MAT标签蛋白,有利于后期将其从酶切体系中去除。
SUMO蛋白酶识别完整的含有100个氨基酸的SUMO(Small Ubiquitin-like Modifier)标签蛋白,并能高效地把SUMO从融合蛋白上切割下来。与EK和TEV等蛋白酶的识别位点相比,由于其识别序列长,所以SUMO蛋白酶酶切反应有很高的特异性,且在较宽范围的反应环境体系中保持较高的活力,例如温度(4-30℃)、pH(5.5-9.5)等。SUMO蛋白酶还具有多聚His标签,便于使用亲和层析的方法去除SUMO蛋白酶,纯化目的蛋白。


|
产品名称 |
货号 |
规格 |
|
UCF.ME® Recombinant Enterokinase(His Tag) 重组肠激酶(His标签) |
100 U/500 U/5000 U/ 100 KU/1 MU(1000 KU) |
|
|
UCF.ME® Recombinant Enterokinase GMP-grade(His Tag) GMP级重组肠激酶(His标签) |
5000 U/100 KU/ 1 MU(1000 KU) |
|
|
Bovine Thrombin,High Specific Activity,>2000 IU/mg 牛凝血酶(高活力,>2000 IU/mg) |
1 KU/2 KU |
|
|
UCF.ME® rTEV Protease (5 U/μL) |
1 KU/10 KU/50 KU/ 1 MU(1000 KU) |
|
|
3C Protease |
100 U/500 U/2×500 U |
|
|
SUMO Protease |
100 U/500 U /1000 U/10 KU |
|
|
HisSep Ni-NTA Agarose Resin His标签蛋白琼脂糖纯化树脂 |
10 mL/50 mL/100 mL |
|
|
Heparin Agarose 6FF 肝素亲和纯化介质 |
5 mL/25 mL/100 mL |
|
|
GSTSep Glutathione Agarose Resin 谷胱甘肽琼脂糖树脂(用于GST标签蛋白纯化) |
10 mL/50 mL/100 mL |



