

 选质粒,还是有点技术含量的
选质粒,还是有点技术含量的
8215
2022-11-17
进行基础研究或应用科学研究,多少会涉及到质粒工具,这些质粒无论是自己构建还是别人馈赠或者公司购买前后,我们要尽可能做到知其然知其所以然,便于知悉获得的质粒是否适用于研究目的。
通过这篇文章,尝试解决:如何快速知道质粒上的各种元件并了解我的质粒是否适用于我的实验,如何通过元件解读质粒的提取、质粒转染甚至宿主中的表达和筛选问题?
一、质粒的工具属性-强大的运载工具
实验研究中的质粒是人为改造的重组质粒,改造之处是以天然质粒或简单质粒骨架为基础量体裁衣式地改变大小、重新改编序列组成。除具备自主复制外,质粒可操作性和功能性更强,更具工具属性。
1、克隆质粒
能在宿主细胞中复制扩增,用于测序和保存外源基因。这类载体具有简单的组成基本元件(ori,Ampr/Kanar,MCS),没有启动子等启动转录、翻译的元件。
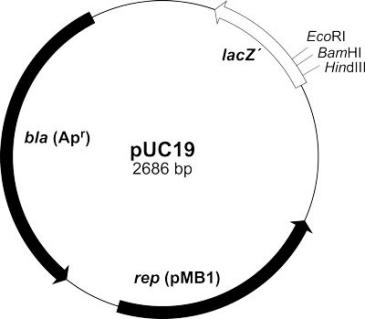
2、基因(过)表达质粒
除克隆载体的基本元件外,还具有启动子等启动转录/翻译所必需的DNA序列。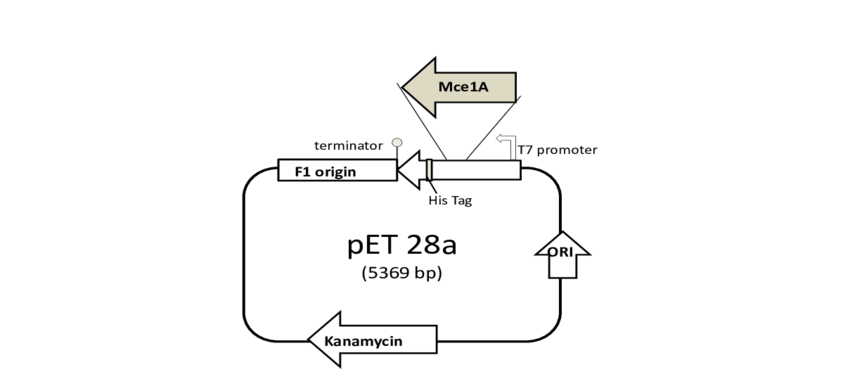
3、基因敲除/敲降质粒
用于靶向基因敲除和定点编辑。含有目标基因的识别序列或目标基因mRNA的shRNA,实现切断目标基因或mRNA降解,起到基因表达沉默的效果,下调蛋白表达。
二、具备上述属性的源头——作用元件的存在
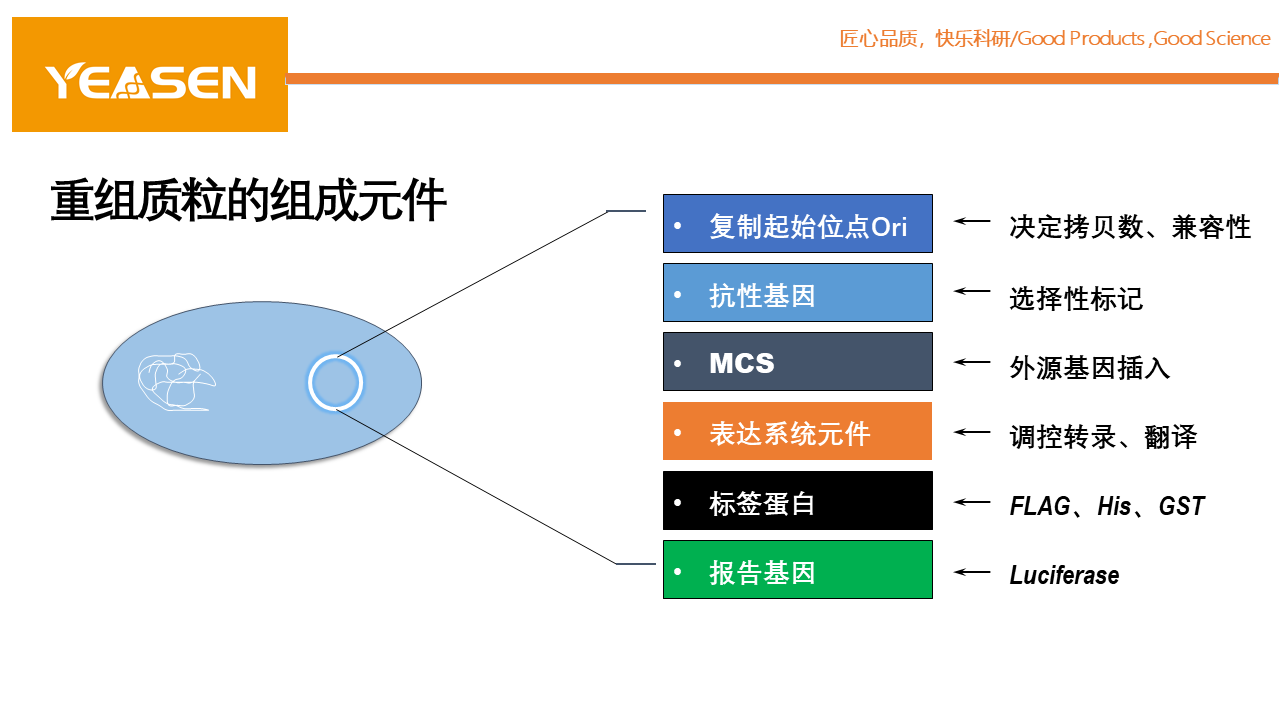
1.复制起始位点Ori
被宿主细胞复制因子所识别,利用宿主的复制机器,复制,增加拷贝数。因宿主复制机器不同,可分为原核/真核/穿梭质粒。
2.抗性基因
质粒转化是一个效率极低的过程,平均约10000个细胞中有1个细胞转化成功。想象一下,如果没有抗性基因的存在,筛选出阳性克隆的时间将大大延长。并且,外源性存在的质粒要进行复制等活动,会消耗宿主细胞自身的资源,在没有抗生素的培养基中,带有质粒的细胞不占优势,慢慢消失;在抗生素的存在情况下,抗性基因帮助宿主细胞分解抗生素,细胞才会需要质粒。
3.多克隆酶切位点
决定能不能放目的基因以及如何放置目的基因。商业化的质粒大多已人为地改造添加多个限制酶的单一切点。便于外源基因的插入。
4.标签蛋白/报告基因
常见的标签如6×His、Myc、GST等;常用到的报告基因如β-半乳糖苷酶、GFP、荧光素酶等。
5.表达系统元件
即启动子(Promoter)-核糖体结合位点(RBS)-转录终止信号-增强子。这是用来区分克隆载体与表达载体的。克隆载体中加入一些表达系统元件即成为表达载体。对于质粒发挥作用,至关重要!
启动子:是RNA聚合酶结合位点,位于基因表达框的上游。启动子序列控制RNA聚合酶与转录因子的结合,因此对于DNA何时何地转录至关重要。因宿主RNA聚合酶种类不同,质粒中的启动子类型也有所不同。
核糖体结合位点:一般为AUG(起始密码子)。也常常会有SD序列,在起始密码子上游5-10bp处,同16s rRNA 3'端互补。
转录终止信号:结构基因的最后一个外显子中有一个AATAAA的保守序列,下游有一段GT或T富丰区,这2部分共同构成poly(A)加尾信号。细菌表达质粒常见的终止子如T7,哺乳动物细胞质粒常见的终止子如SV40、SV40 late polyA、BGH等。
增强子:为真核基因组(包括真核病毒基因组)中的一种具有增强邻近基因转录过程的调控顺序。其作用与增强子所在的位置或方向无关。
三、质粒的选择——如何根据实验目的,选择合适的质粒
对作用元件做了简单的介绍后,我们进一步从实验角度理解这些元件如何服务于我们的实验。质粒选择前,我们先想到的是看质粒图谱,其实也就是思考选择作用元件的过程。
1.元件的选择
第一步,根据实验目的区别选择克隆载体还是表达载体。
第二步,看懂复制起始位点(Ori),Ori是决定质粒拷贝数和宿主类型的重要参数。
l 控制质粒拷贝数:Ori对于复制的控制,可分为松弛型和严谨型。松弛型质粒在细菌内的复制不受控制,拷贝数>10个拷贝,严谨型质粒<10个拷贝。常见的Ori如ColE1和pBR322等,属于松弛型。
l 决定质粒共转染:相同类型的Ori质粒在同一个宿主细胞内不兼容。相同的Ori质粒使用相同的复制机器,易引起宿主细胞环境的不稳定。因此,两质粒或多质粒共转染时,需注意质粒的Ori是否兼容。
l 决定在何种宿主内存在:根据Ori类型,可以将载体分为原核质粒、真核质粒和穿梭质粒。其中穿梭质粒含原核和真核生物2个复制子,以确保两类细胞中都能扩增。
第三步,筛选抗生素的选择。可参考宿主类型,下面列举一些常见的抗性筛选基因。
|
抗性基因 |
常见宿主类型 |
抗性基因作用机制 |
筛选抗生素 |
抗生素作用机制 |
|
AmpR |
大肠杆菌 |
β-内酰胺酶,水解β-内酰胺环,解除氨苄的毒性 |
氨苄青霉素(或羧苄青霉素) |
抑制细胞壁合成 |
|
KanR
|
大肠杆菌 植物 |
氨基糖苷磷酸转移酶使卡那霉素失活 |
硫酸卡那霉素
|
与细菌核糖体30S亚单位结合,抑制细菌蛋白质合成 |
|
TetR |
革兰氏阴性和阳性细菌 |
可以阻止四环素进入细胞 |
四环素盐酸盐 |
与核糖体30S亚单位的结合来阻止蛋白质的合成 |
|
NeoR
|
细菌、酵母、动植物细胞 |
氨基糖苷磷酸转移酶抑制抗生素-核糖体间的相互作用,从而使得抗生素失活 |
G418
|
与核糖体80S亚单位结合,抑制蛋白质合成 |
|
NeoR
|
细菌、酵母、动植物细胞 |
氨基糖苷磷酸转移酶抑制抗生素-核糖体间的相互作用,从而使得抗生素失活 |
G418
|
与核糖体80S亚单位结合,抑制蛋白质合成 |
|
PuroR |
革兰氏阳性菌 动物细胞
|
乙酰化嘌呤霉素使其失活 |
嘌呤霉素 |
核糖体A位点结合,终止蛋白质合成 |
|
HygR |
各类原核、真核、哺乳动物细胞
|
将潮霉素B转化成不具有生物活性的磷酸化产物 |
潮霉素B |
干扰80S核糖体易位和诱导对mRNA模板的错读而抑制蛋白合成 |
第四步,酶切位点的选择。如果进行蛋白表达,注意避免阅读框移位。
第五步,注意选择合适的标签蛋白或标记基因。
第六步,读懂表达作用元件,最大程度激发质粒功能!质粒上携带的各类基因唯有表达为蛋白,才能发挥作用。1)启动子方向性:我们看质粒图谱,会发现顺时针和逆时针两类箭头,这是基因表达的方向。启动子是在靶基因的上游位置。2)启动子特异性:真核生物常见的启动子如CMV、EF1α、SV40,原核生物常见的如T7启动子。
2.几点注意事项
关于宿主与启动子的选择:必须搭配对应的启动子,方能启动基因的表达。如neo抗性基因可以消除G418、卡那霉素的作用,在pET-28a质粒中,neo基因的启动子可以在大肠杆菌启动抗性基因的表达,用于卡那霉素的筛选,但不能在真核生物中用于G418的筛选;在酵母载体pPIC9K中,携带的neo基因可以用于大肠杆菌卡那霉素和酵母G418筛选。
关于质粒载量:复制子复制能力有限,插入片段越大,会造成质粒不稳定,同时复制困难。
关于拷贝数:并不是越多越好。质粒复制借助于宿主,拷贝数越多,宿主消耗越严重,对宿主影响越大。



