

 基因编辑在免疫细胞治疗中的应用
基因编辑在免疫细胞治疗中的应用
10647
2024-07-02
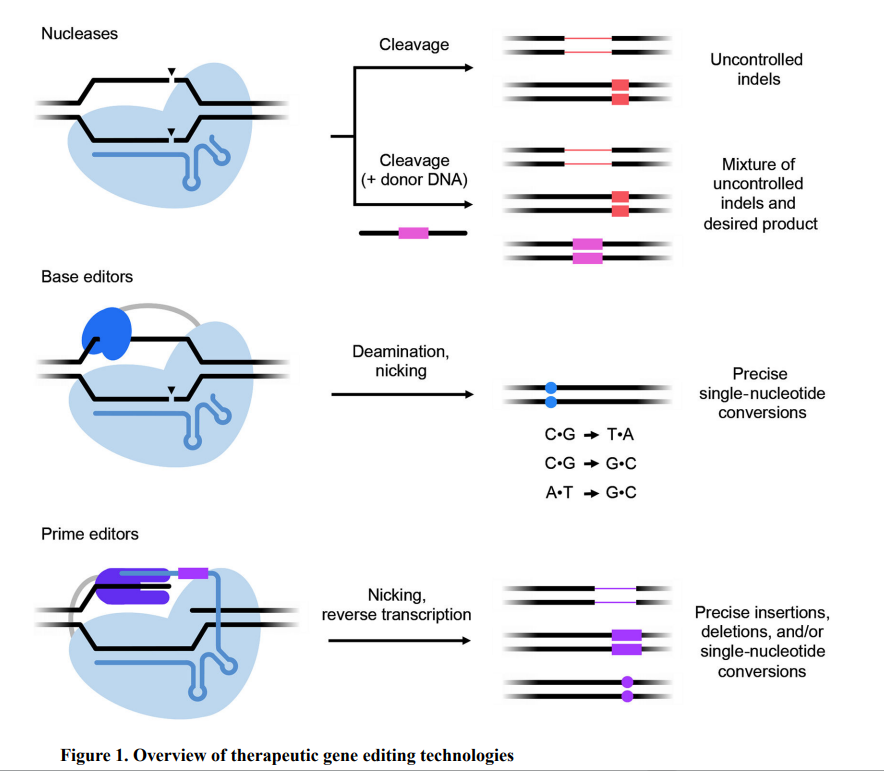
图1.基因编辑技术分类[1]
可编程核酸酶主要包括锌指核酸酶 (ZFNs) 和转录激活因子样效应核酸酶 (TALENs) 以及CRISPR-Cas核酸酶,这三种编辑方式都是将目的DNA打断后产生双链断裂(DSB),后续依赖非同源末端连接或同源定向重组的方式进行修复。ZFNs是最早用于哺乳动物细胞基因编辑的人工核酸酶,通过锌指结构域识别DNA序列并通过Fok1的二聚体对DNA进行定点切割。ZFNs作为第一代基因编辑技术其基因修复方式多样,对基因表达程度影响较小,但是其设计与筛选过程复杂,脱靶风险高并且细胞毒性大。TALENs作为第二代基因编辑技术,结构类似于ZFNs,其毒性较低且构建容易。然而,TALENs在尺寸上比ZFNs大,组装更加困难。后续出现的CRISPR-Cas核酸酶的出现则开启了新的基因编辑时代。相较于ZFNs和TALENs,CRISPR/Cas具有良好的柔性和高效性,可以同时敲除多个基因位点,其中以CRISPR-Cas9应用最为广泛。CRISPR/Cas9基因编辑系统包括Cas9蛋白和单链向导 RNA (sgRNA),在sgRNA的向导下通过碱基互补配对原则从而识别目的基因序列,并引导Cas9蛋白酶有效切割DNA双链,形成双链断裂损伤后修复。
BE系统主要由dCas9,sgDNA和脱氨酶组成,通过对基因组 DNA中的单核苷酸转换以实现精确的基因校正,无需产生DSB,从而克服了核酸酶的许多限制,主要包括胞嘧啶碱基编辑器 (CBE)和腺嘌呤碱基编辑器 (ABE)。PE则进一步在BE的系统引入逆转录酶,将切口酶与逆转录酶偶联在一起,实现单碱基的转换,敲除以及添加等操作。碱基编辑技术具有编辑效率高、编辑损伤少等特点,但是脱氨酶的使用可能会导致癌变风险的增加,而且可能出现的脱靶效应会严重影响编辑效率,其可靠性有待进一步研究。
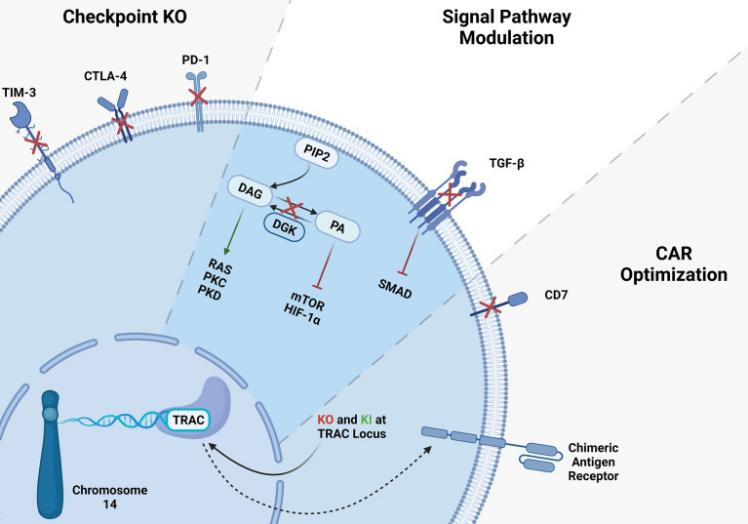
图2.CRISPR技术在T细胞治疗中的应用[2]
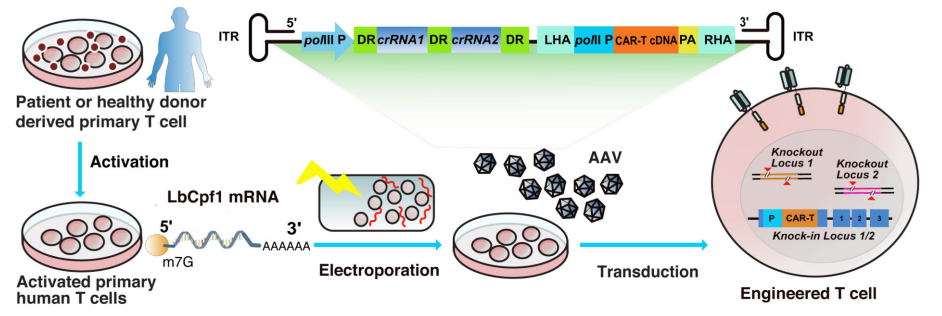
图3.AAV-cpf1介导的多重基因编辑流程[5]
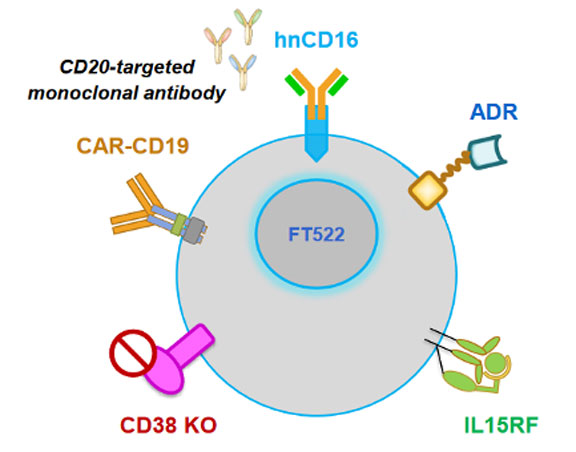
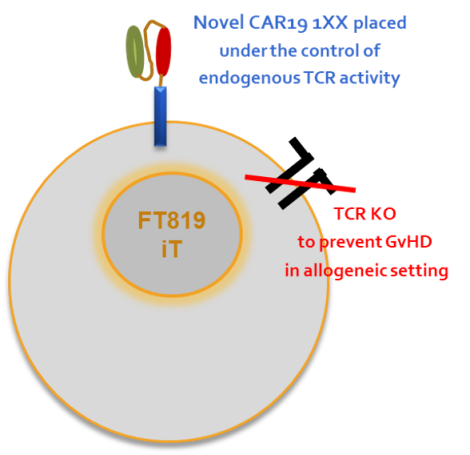
图4 . FT819结构 图5 . FT522结构
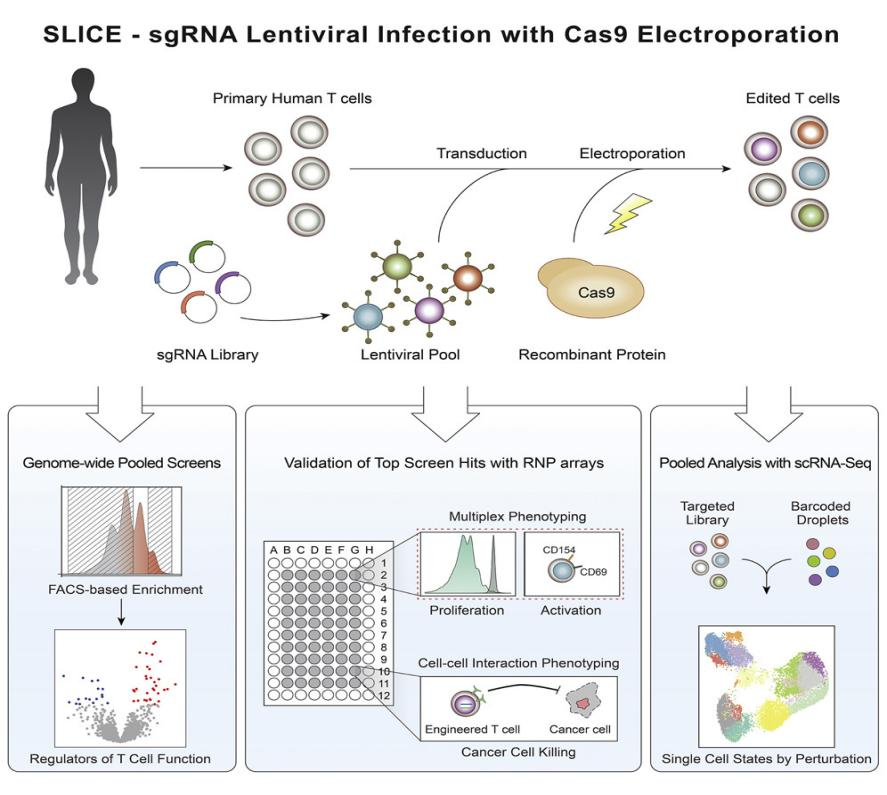
图6.SLICE筛选平台[8]
首先,对于基因编辑策略而言, 最大的担忧是潜在的脱靶毒性,一旦发生脱靶,将导致非靶标基因的改变或调控元件的破坏,会造成不可逆转的后果。尽管利用预测软件能够分析出潜在脱靶位点, 但是在一些非典型PAM位点或者与靶点相似度较低位点仍然有可能有脱靶现象发生。同时,全基因组测序分析对于概率极低的脱靶现象也很有可能出现假阴性的结果。近年来, 许多灵敏度极高的体内、体外实验方法被开发出来例如GUIDE-Seq, CIRCLE-seq, CHANGE-Seq等以帮助检测潜在的低频脱靶位点。新型基因编辑系统例如PE以及BE编辑系统也逐渐应用于免疫细胞治疗中。这些方法在免疫细胞治疗中的合理运用将会对临床实验的安全性起到重要的作用。
同时以过继性细胞治疗为基础的肿瘤免疫细胞治疗,主要包括CAR-T细胞、TCR-T细胞和CAR-NK细胞治疗等,均需要足够数量的免疫细胞保证其临床疗效。目前大多数基于CRISPR/Cas技术编辑的T细胞都是通过电穿孔的方法进行转导,但是该方法对细胞损伤较大,致使细胞的活力下降并阻碍其体外增殖。因此,其他更安全、更高效的传递途径亟待探索。
1.Raguram A. et al. Therapeutic in vivo delivery of gene editing agents. Cell. 2022 Jul 21;185(15):2806-2827.
2.Elmas E. et al. CRISPR Gene Editing of Human Primary NK and T Cells for Cancer Immunotherapy. Front Oncol. 2022 Apr 5;12:834002.
3.Stadtmauer EA. et al. CRISPR-engineered T cells in patients with refractory cancer. Science. 2020 Feb 28;367(6481):eaba7365.
4.J. Eyquem. et al., Targeting a CAR to the TRAC locus with CRISPR/Cas9 enhances tumour rejection, Nature 543 (7643) (2017) 113–117.
5.Dai X. et al. One-step generation of modular CAR-T cells with AAV-Cpf1. Nat Methods. 2019 Mar;16(3):247-254.
6.Dai X. et al.. Massively parallel knock-in engineering of human T cells. Nat Biotechnol. 2023 Jan 26. doi: 10.1038/s41587-022-01639-x.
7.Zhu H. et al. Metabolic Reprograming via Deletion of CISH in Human iPSC-Derived NK Cells Promotes In Vivo Persistence and Enhances Anti-tumor Activity. Cell Stem Cell. 2020 Aug 6;27(2):224-237.
8.Shifrut E. et al.Genome-wide CRISPR Screens in Primary Human T Cells Reveal Key Regulators of Immune Function. Cell. 2018 Dec 13;175(7):1958-1971.e15.
9.Wang D. et al. CRISPR Screening of CAR T Cells and Cancer Stem Cells Reveals Critical Dependencies for Cell-Based Therapies. Cancer Discov. 2021 May;11(5):1192-1211.
10.Parola C. et al. Antibody discovery and engineering by enhanced CRISPR-Cas9 integration of variable gene cassette libraries in mammalian cells. MAbs. 2019 Nov-Dec;11(8):1367-1380.



