

 什么是铁死亡(Ferroptosis)?铁死亡的文献分析和相关实验检测试剂汇总
什么是铁死亡(Ferroptosis)?铁死亡的文献分析和相关实验检测试剂汇总
15567
2024-06-17
铁死亡的背景和特点
铁死亡(Ferroptosis)是一种由铁依赖的脂质过氧化驱动的细胞死亡形式,十年前被发现,并因此命名。铁死亡涉及到广泛的生物学环境,从发展到衰老、免疫和癌症。铁死亡不同于细胞凋亡和其他形式的细胞死亡,铁死亡的发现出现在2003年的小分子诱导非凋亡形式的细胞死亡中。
由铁依赖的细胞ROS的积累导致细胞氧化还原稳态的破坏而导致的细胞死亡被称为铁死亡。ROS的积累超过了谷胱甘肽 (GSH) 和谷胱甘肽过氧化物酶4(GPX4)的氧化还原含量。
铁死亡和细胞凋亡、细胞焦亡、PARP1依赖性细胞死亡、细胞坏死的区别,见下图汇总。[2]
|
细胞死亡类型 |
形态学特征 |
调节因子 |
生化特征 |
ROS功能 |
|
铁死亡 |
细胞膜未发生破裂或起泡;线粒体嵴减少或消失;线粒体外膜破裂;细胞核大小正常;染色质正常 |
NOX, GPX4, DHODH, FSP1, BH4, GOT1, NRF2 |
脂质、铁和ROS促进铁死亡; GPX4、FSP1和DHODH协同抑制铁死亡 |
促进脂质过氧化,进一步诱导4-hydroxy-2-nonanalproduction生成,导致细胞膜破裂和铁死亡 |
|
细胞凋亡 |
质膜起泡;细胞和核体积减少;核碎裂;染色质缩合 |
p53, Bax, Bak, Bcl-2, Bcl-XL |
DNA断裂;半胱天冬酶激活;磷脂酰丝氨酸转移 |
过量的ROS可诱导自噬,并通过降解受损的线粒体来维持细胞内稳态 |
|
细胞焦亡 |
细胞膨胀形成突起;细胞膜形成孔隙;细胞核收缩;DNA断裂 |
CASP1, CASP4, CASP5, GSDMD |
激活caspase-1;促进炎症细胞因子IL-1β和IL-18产生;GSDMD是细胞焦亡的关键执行者 |
ROS激活经典焦亡途径,依赖于NLRP 3炎症小体来激活Caspase-1 |
|
Parthanatos(PARP1依赖性细胞死亡) |
染色质凝结;产生大量DNA片段; |
PARP1, AIFM1 |
PARP1激活产生大量独立于caspases的PAR |
过量的ROS引发细胞程序性死亡 |
|
细胞坏死 |
质膜破裂;细胞器肿胀;中度染色质凝结 |
RIP1/3, MLKL |
RIP1、RIP3和MLKL PARP1的过度激活 |
RIPK1自磷酸化,RIPK3募集和坏死体形成 |
铁死亡的机制
新陈代谢产生了铁死亡的底物和天然抑制剂;
脂质代谢: ACSL4和LPCAT3产生铁死亡的脂质驱动因素。铁死亡最终是由特定的膜脂质的过氧化作用驱动的;
GPX4独立控制脂质ROS积累。GPX4是铁死亡的核心抑制剂。在随后的几十年里,它被证明在线粒体的能量生产中具有重要的电子传递作用。
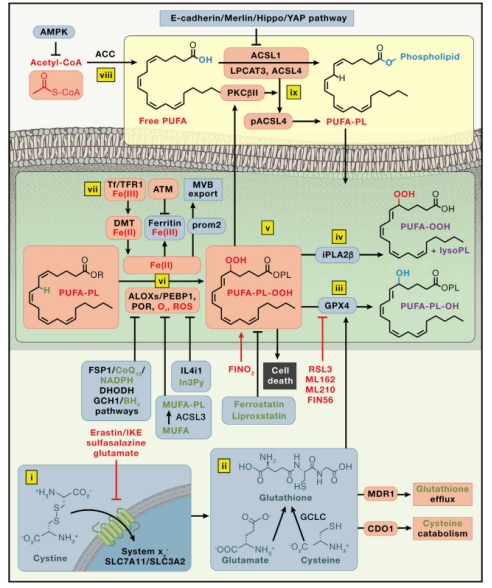
铁死亡机制
(蛋白质用黑色标注;诱导铁死亡的用红色框标注;抑制铁死亡的用蓝色框标注;其中绿色标注的为抑制铁死亡的小分子化合物、脂质等物质;红色标注的为诱导铁死亡的物质;紫色标注的为既不抑制又不诱导铁死亡的物质)
图片来源于文献(PMID: 35803244)[1]
抗铁死亡的机制
抗铁死亡的极致包括代谢、ROS生物学和铁的调节:
至少有三种主要的铁死亡抗性机制已经被确定——抗氧化调节因子NRF2、转导途径和雷帕霉素(mTOR)的机制靶点。NRF2是抗氧化反应的主调节因子,它驱动抑制铁死亡的转录反应。NRF2通过激活微粒体GSH s-转移酶1 (MGST1)来抑制铁死亡,从而抑制GSH。[1]
1. 转硫途径
转硫途径通过从蛋氨酸中产生半胱氨酸(cys),克服cys饥饿产生的抑制系统xc来驱动对铁死亡的抗性。 2016年对铁死亡抑制因子进行的siRNA筛选显示,最热门的是半胱氨酸-tRNA合成酶,通过激活转硫途径抑制Erastin诱导的铁死亡。
2. mTOR途径
mTOR途径通过增加GPX4蛋白合成和增加固醇反应元件结合蛋白(SREBP)介导的脂肪生成来促进对铁死亡的抗性。因此,铁死亡抑制可以通过控制铁稳态、ROS丰度和代谢来产生。
3. 驱动铁死亡的亚细胞和细胞器
铁死亡是通过过氧化物脂质的积累来完成的。近年来,人们探索了含有这些氧化脂质的亚细胞膜,阐明了不同细胞器对铁死亡的贡献
4. 质膜破裂是铁死亡的晚期事件
铁死亡的最后阶段涉及质膜的通透性。铁死亡向邻近细胞的传播可能依赖于具有完整质膜的氧化脂质的释放,可能通过胞外囊泡;释放的氧化脂质可能在邻近细胞中引发铁死亡。
5. 内质网是铁死亡过程中脂质过氧化的重要部位
抑制内质网中的脂质过氧化足以遏制铁死亡,通过影响脂质过氧化来调节铁死亡的化合物主要定位于内质网,这表明内质网是铁死亡过程中脂质过氧化的最关键部位。
6. 线粒体可以作为半胱氨酸(cys)饥饿诱导的铁死亡的启动子和放大器
一些研究表明,线粒体可以作为由cys饥饿和GSH耗尽引起的铁死亡的启动器或放大器。线粒体DHODH和线粒体CoQ10以这种方式触发铁死亡。抑制线粒体电子传递链也可以减弱cys饥饿引起的铁死亡以及线粒体的损耗。
7. 过氧化物酶体可以通过醚脂质的过氧化作用来驱动铁死亡
虽然过氧化物酶体ROS在铁死亡中尚未直接检测,但可能过氧化物酶体产生的H2O2可能引发脂质过氧化,可能与过氧化物酶体铁(II)有关。
8. 溶酶体是铁的储存库,可引发铁死亡
铁抑制素在溶酶体中积累,但它们在溶酶体的局部化并没有助于它们抑制Erastin或RSL3诱导的铁死亡。因此,在这种情况下,溶酶体通常不会导致铁死亡。然而,溶酶体是铁的储存库,原则上可以引发铁死亡。
9. 高尔基体应力可促进铁死亡
在某些情况下,用brefeldin A(布雷非得菌素A,BFA)等药物可触发高尔基体应力,引发铁死亡。
总之,内质网是驱动脂质过氧化的中心枢纽,脂质过氧化作用通过铁死亡促进细胞死亡。这是合乎逻辑的,因为大量的脂质和多不饱和脂肪酸代谢(PUFA)代谢发生在ER中。其他细胞器,如线粒体、溶酶体、过氧化物酶体和高尔基体,可以通过启动脂质过氧化或增强ROS的产生来启动或增强细胞对铁死亡的敏感性。最终,脂质过氧化物扩散到质膜,引发质膜破裂。
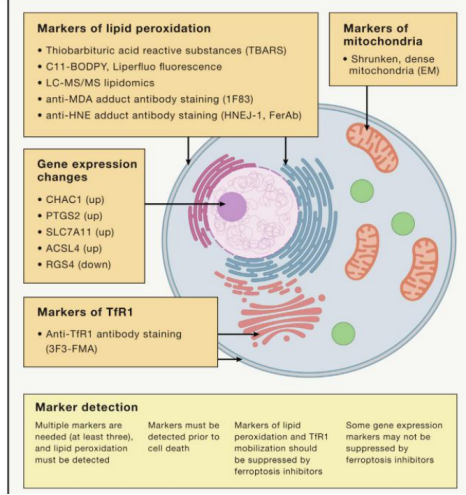
铁死亡标志物
图片来自于文献(PMID: 35803244)[1]
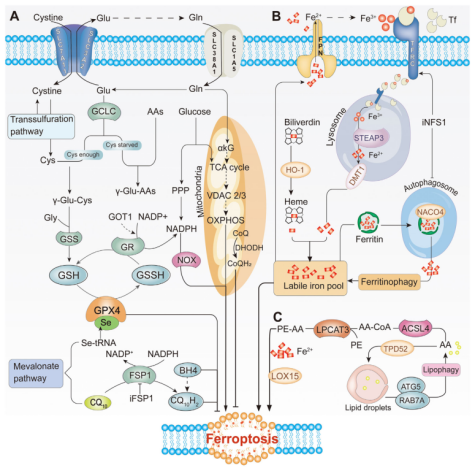
铁死亡的代谢途径
图片来源于(PMID: 35133083)[2]
铁死亡诱导剂
|
诱导剂 |
货号 |
作用通路 |
|
54037ES |
抑制system Xc到GSH的缺乏 |
|
|
53274ES |
抑制system Xc到GSH的缺乏 |
|
|
53468ES |
抑制system Xc到GSH的缺乏 |
|
|
53469ES |
抑制GPX4到脂质过氧化 |
|
|
54038ES |
抑制GCS到GSH的缺乏 |
|
|
54039ES |
消耗CoQ10,降低GPX4水平 |
|
|
54040ES |
引发脂质物氧化,间接抑制GPX4 |
铁死亡抑制剂
|
抑制剂 |
货号 |
作用通路 |
|
54020ES |
阻断脂质物氧化 |
|
|
54021ES |
阻断脂质物氧化 |
|
|
50304ES |
阻断脂质过氧化物的堆积 |
|
|
54023ES |
阻断脂质过氧化物的堆积 |
|
|
54024ES |
阻断脂质过氧化物的堆积 |
|
|
54025ES |
阻断脂质物氧化 |
|
|
54026ES |
阻断脂质物氧化 |
|
|
54027ES |
螯合铁离子 |
|
|
54028ES |
螯合铁离子 |
|
|
54029ES |
螯合铁离子 |
|
|
54030ES |
螯合铁离子 |
|
|
54031ES |
抗氧化作用:脂质体修复 |
|
|
54032ES |
阻断脂质物氧化 |
|
|
54033ES |
阻断脂氧合酶诱导的脂质过氧化 |
|
|
54034ES |
阻断脂氧合酶诱导的脂质过氧化 |
|
|
54035ES |
阻断脂氧合酶诱导的脂质过氧化 |
|
|
54036ES |
阻断脂氧合酶诱导的脂质过氧化 |
铁死亡检测试剂盒
|
产品名称 |
货号 |
作用通路 |
|
50101ES |
高灵敏度活性氧检测试剂盒 |
|
|
GSH/GSSG Ratio Fluorimetric Detection Assay Kit (Green Fluorescence) GSH/GSSG比例荧光检测试剂盒(绿色荧光 |
50120ES |
高灵敏度GSH/GSSG定量试剂盒(荧光法) |
|
NAD/NADH Ratio Fluorimetric Detection Assay Kit (Red Fluorescence) NAD/NADH比率荧光检测试剂盒(红色荧光) |
50130ES |
高灵敏度NAD/NADH定量试剂盒(荧光法 |
|
NADP/NADPH Ratio Fluorimetric Detection Assay Kit (Red Fluorescence) NADP/NADPH比率荧光检测试剂盒(红色荧光) |
50140ES |
高灵敏度NADP/NADPH定量试剂盒(荧光法) |
|
60408ES |
葡萄糖检测试剂盒 |
|
|
60533ES |
检测灵敏度极高的一种铁染色方法 |



