
国际期刊|Cell!十字花科植物一年生转变为多年生关键基因揭秘!
你是否曾想过,为什么有些植物一年只开一次花,而有些植物却可以年年开花?这背后,其实有着复杂的遗传机制。
在植物学中,一年生植物会在一年的周期中经历发芽、生长、开花、结果,直至死亡。比如大豆、玉米、土豆等作物。而多年生植物在开花、结果、枝叶老去之后,在来年依旧能萌发新芽,循环往复,历经数年,如冬小麦、白菜、胡萝卜等。一年生与多年生植物之间并不存在着一道界线分明的分水岭。比如,红薯、鼠曲草在日本属于一年生草本植物,而在热带地区它们却成了多年生植物。人们对于转变背后的进化模式和遗传基础仍然知之甚少。
在追求农业可持续发展的今天,多年生作物的重要性不言而喻。它们不仅能够减少农业劳动力的投入,降低生产成本,还能对环境保护产生积极影响。但是多年生作物的繁殖难题一直是农业领域的挑战。那么,到底是什么遗传机制调控一年生转变为多年生植物呢?我们能否解析这种转变机制,进而调控植物的生长周期,为农业的可持续发展出一份力,实现年年丰收的愿景呢?
2024年5月28日,国际顶级期刊Cell在线发表了中国科学院分子植物科学卓越创新中心王佳伟团队的最新研究:“Reciprocal conversion between annual and polycarpic perennial flowering behavior in the Brassicaceae”。该研究揭示了十字花科植物中,一年生和多果多年生开花行为的转变机制,并利用基因编辑技术成功将一年生植物变成多年生植物,为培育新型多年生作物提供了理论基础。
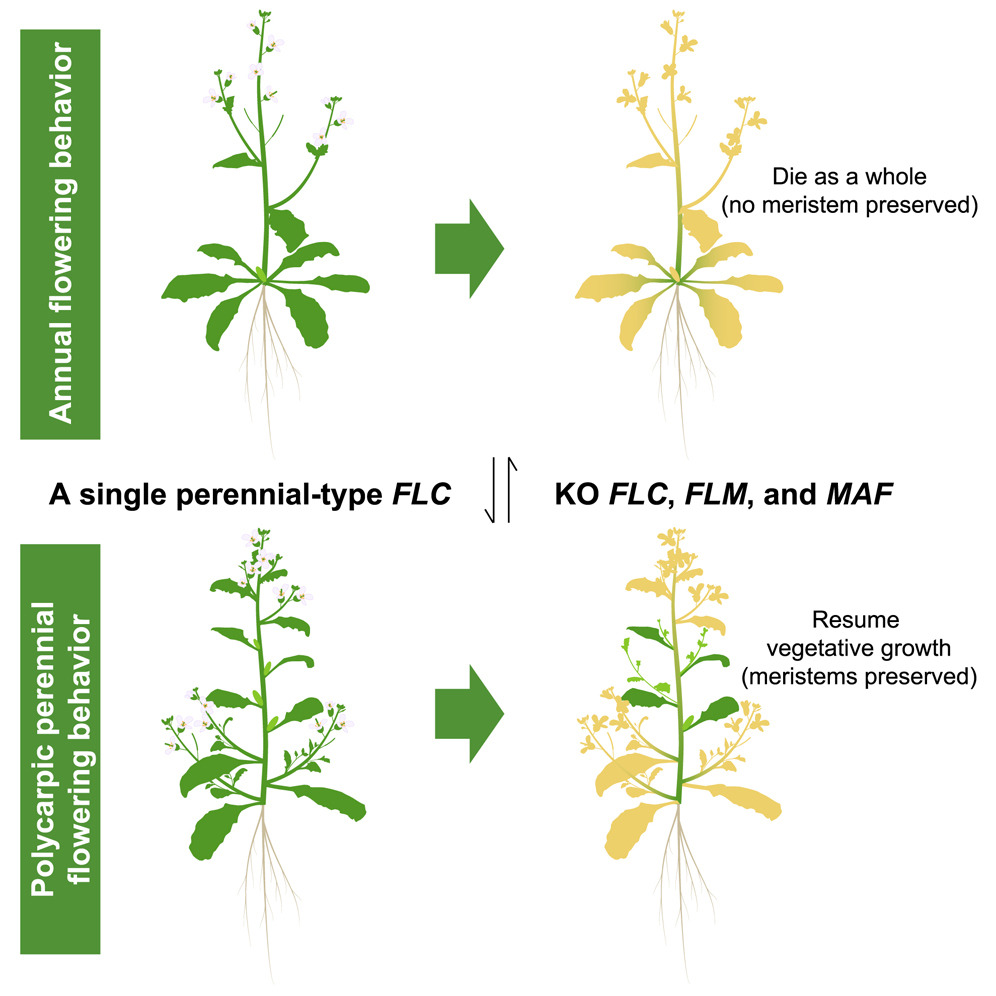
图1. 一年生与多年生植物转化模型(图片来自原文)
研究人员首先对十字花科的所有属进行了筛选比较,获得了两个二倍体多果多年生物种作为模型进行研究,分别是来自SENDAI拟南芥种子储备中心(SASSC)的喜马拉雅须弥芥(Crucihimalaya himalaica)和来自USDA-ARS种质资源信息网络(GRIN)的内华达糖芥(E. nevadense),喜马拉雅须弥芥生长在青藏高原,是研究极端环境下物种形成和生态适应的新模型系统,而内华达糖芥是内华达山脉山峰的特有种。对他们在生长室条件下的生长习性进行表征,E. nevadense在长日照(LDs)中表现出与C. himalaica相当的开花表型(图2),且经过两个月的冷处理之后可以进行新一轮的开花。表明在研究人员的生长室条件下,喜马拉雅须弥芥和内华达糖芥都是典型的多年生多果植物。

图2. 两种多果多年生模型的表征(图片来自原文)
为了进行系统的遗传学研究,研究人员又筛选出了与喜马拉雅须弥芥和内华达糖芥同属的植物,一次结果一年生植物小花糖芥(Erysimum cheiranthoides)和呈现出兼性冬性一年生/多次结实多年生特征的卵叶须弥芥(Crucihimalaya wallichii)。
在此基础上,研究团队利用基因组测序技术对四种植物的基因组进行了测序和组装,获得了这些植物的基因组信息,并通过基因定位技术确定了两个属中负责开花行为自然变异的基因。如图3,在须弥芥属和糖芥属中都成功定位到了三个基因区间(FLC、FLM和MAF)。通过比对模式生物拟南芥的基因功能注释,结果显示这三个遗传区间内存在一类亲缘关系密切的MADS-box转录因子编码基因,它们都具有抑制植物开花(生殖生长)的功能。
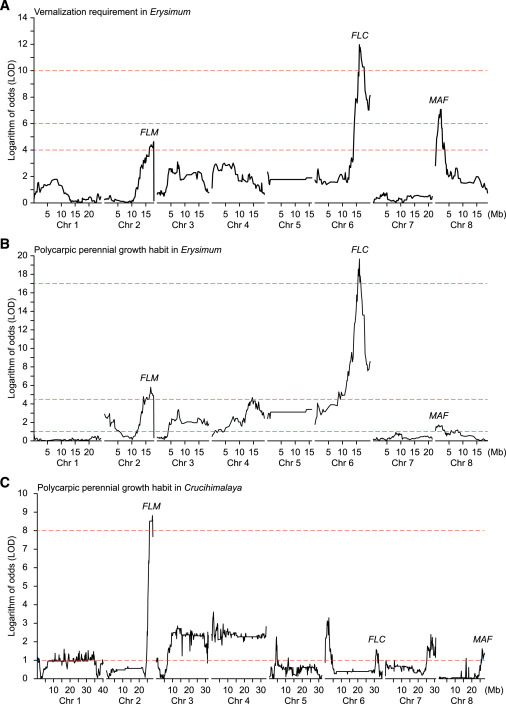
图3. 春化需求和多年生多果植物生长习性的遗传图谱(图片来自原文)
研究人员利用基因克隆和表达分析技术,研究了三种基因(FLC、FLM和MAF)在植物开花行为中的作用。首先,通过CRISPR-Cas9基因编辑技术生成FLC、FLM和MAF基因的功能缺失突变体,即在编码区内或两侧设计两个单向导RNA(sgRNAs),创建了具有大片段缺失的突变体。在喜马拉雅须弥芥中获得了FLC、FLM和MAF的单一突变体,分别将其命名为Chflc、Chflm和Chmaf。
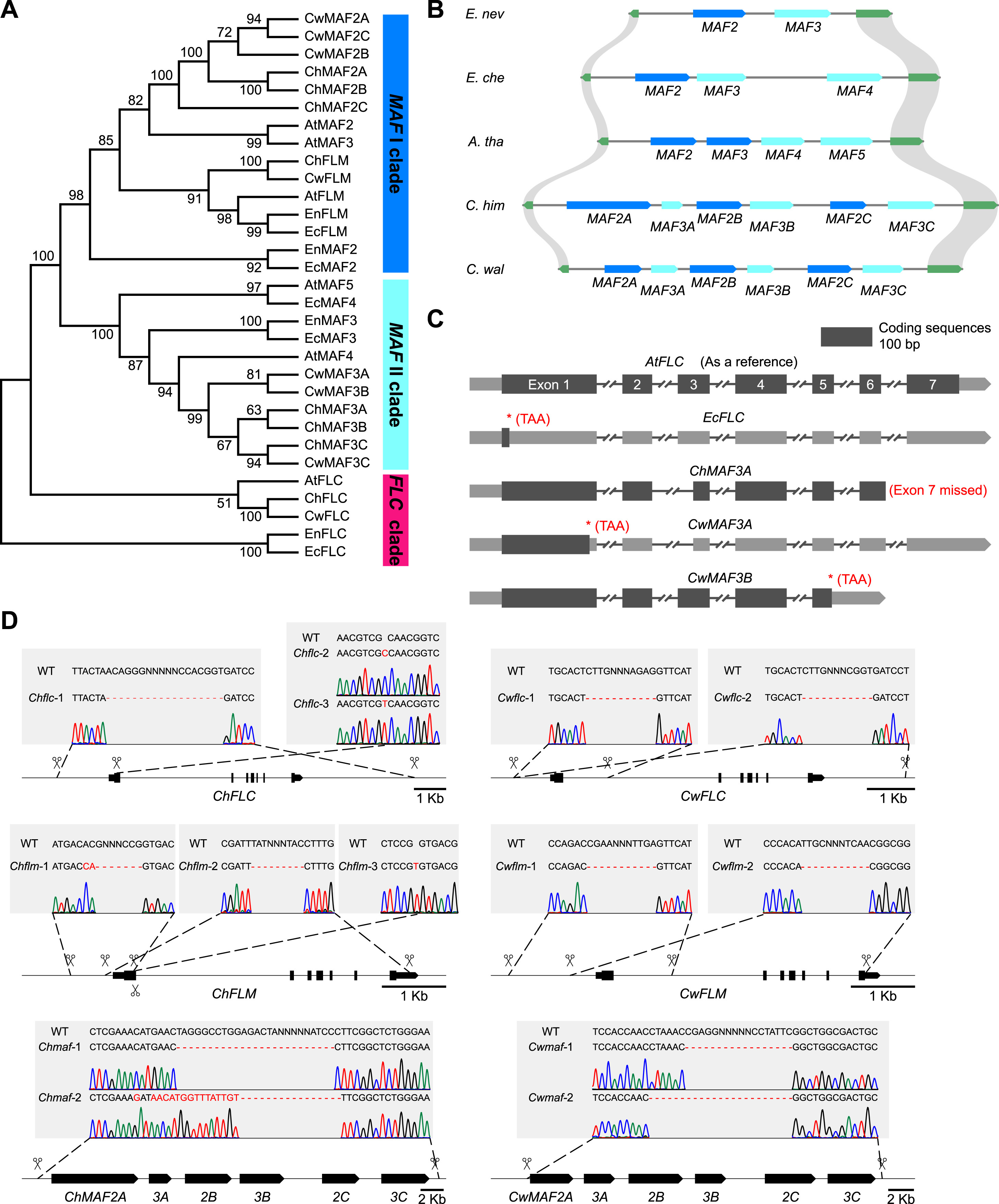
图4. 须弥芥属和糖芥属中类FLC MADS-box基因突变体的生成(图片来自原文)
其次,观察突变体的表型变化。3个突变体在春化之后都可以恢复营养生长(即形成新的营养芽,如图5A所示),都显示出了对于春化的需求。与Chflc和Chflm突变株相比,Chmaf需要更长时间的冷处理(18周)才能完全饱和春化需求。结果显示,FLC、FLM和MAF基因之间存在功能冗余,其中任何一种基因的突变都会影响植物的开花时间,但不同基因的影响程度不同。
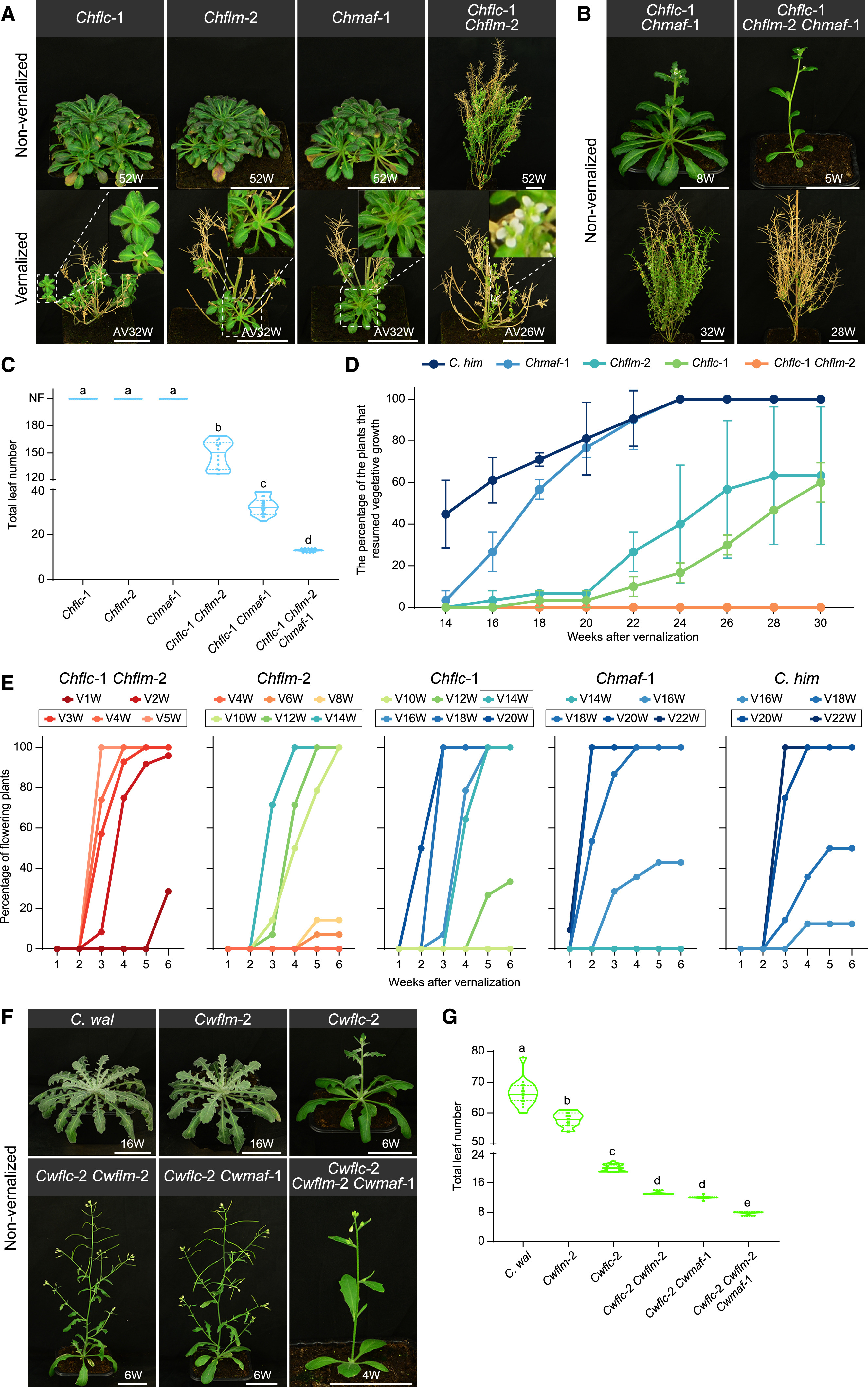
图5. 喜马拉雅须弥芥和卵叶须弥芥类FLC的MADS-box基因突变体的开花时间和胎次(图片来自原文)
接着,通过杂交手段将突变基因以不同的方式组合,成功再现了植物从多次结实多年生逐渐演化为二年生再转变为一年生的轨迹:当FLC、FLM和MAF三个基因都保持功能完好时,植物表现出稳健的多次结实多年生表型。缺失其中一个或两个基因会导致植物出现兼性生活史策略,Chflc Chflm双突变体表现出冬季一年生表型,在LDs中产生150片叶子后开花(图5C),春化可以加速开花(图5E),即从多年生植物转变为了二年生植物;Chflc Chflm突变体显示出了一年生表型,当在LDs中连续生长时,Chflc Chflm植物最终可以永久开花(图5A)。而当三个基因完全缺失时,Chflc Chflm Chmaf三突变体植物转变为一年生植物(图5B和5C)。这些发现与绘图(图1)结果一致,并表明以下几点:
ChFLC、ChFLM和ChMAF对春化的调控存在功能冗余;
ChFLC和ChFLM的共同作用确保了喜马拉雅多果多年生生长习性;
ChMAF强烈抑制开花,主要在春化中发挥作用。
为了更深入地研究这些基因的表达调控机制,研究人员利用染色质免疫沉淀测序(ChIP-seq)和RNA测序(RNA-seq)技术,研究了类FLC基因与染色质结合的位点和下游靶基因的表达变化。揭示了从多果多年生到二年和一年生开花行为的转变是由三个密切相关的MADS-box基因的剂量决定的连续体。多年生植物中,类FLC基因在低温处理后表达量会降低,随后恢复,这与多年生植物可以多次开花的特性相符。这些基因的表达模式、功能强度和组合的多样化赋予了物种采用各种生活史策略的潜力。
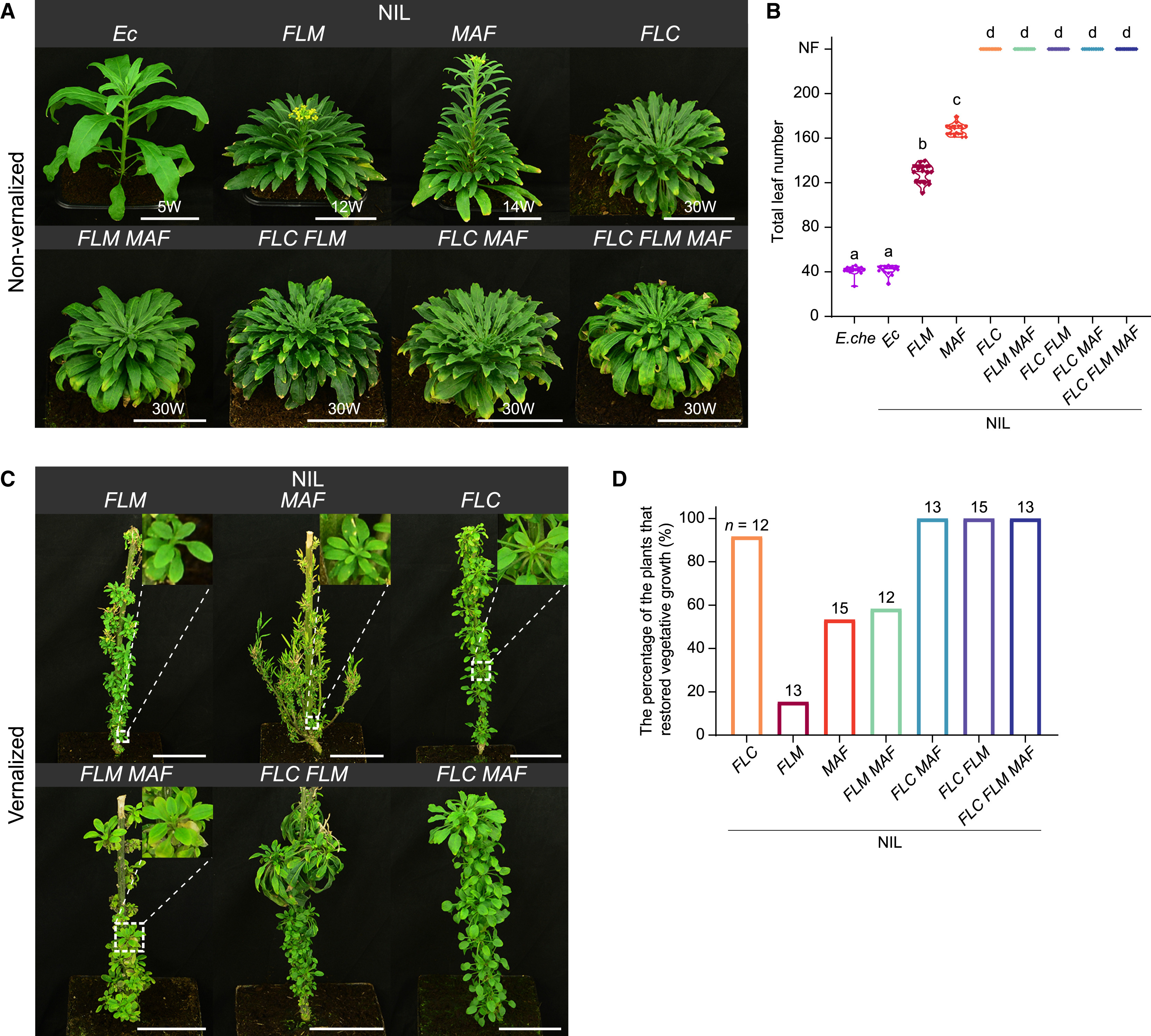
图6. 小花糖芥中NILs的开花时间和胎次(图片来自原文)
最后,研究人员利用转基因技术,尝试培育了一种多果多年生开花甘蓝型油菜植物。通过构建转基因植物,实现多年生类FLC基因的高表达水平,如图7,在春化后可以恢复营养生长,这些一年生植物变成了多年生植物,可以多次开花。
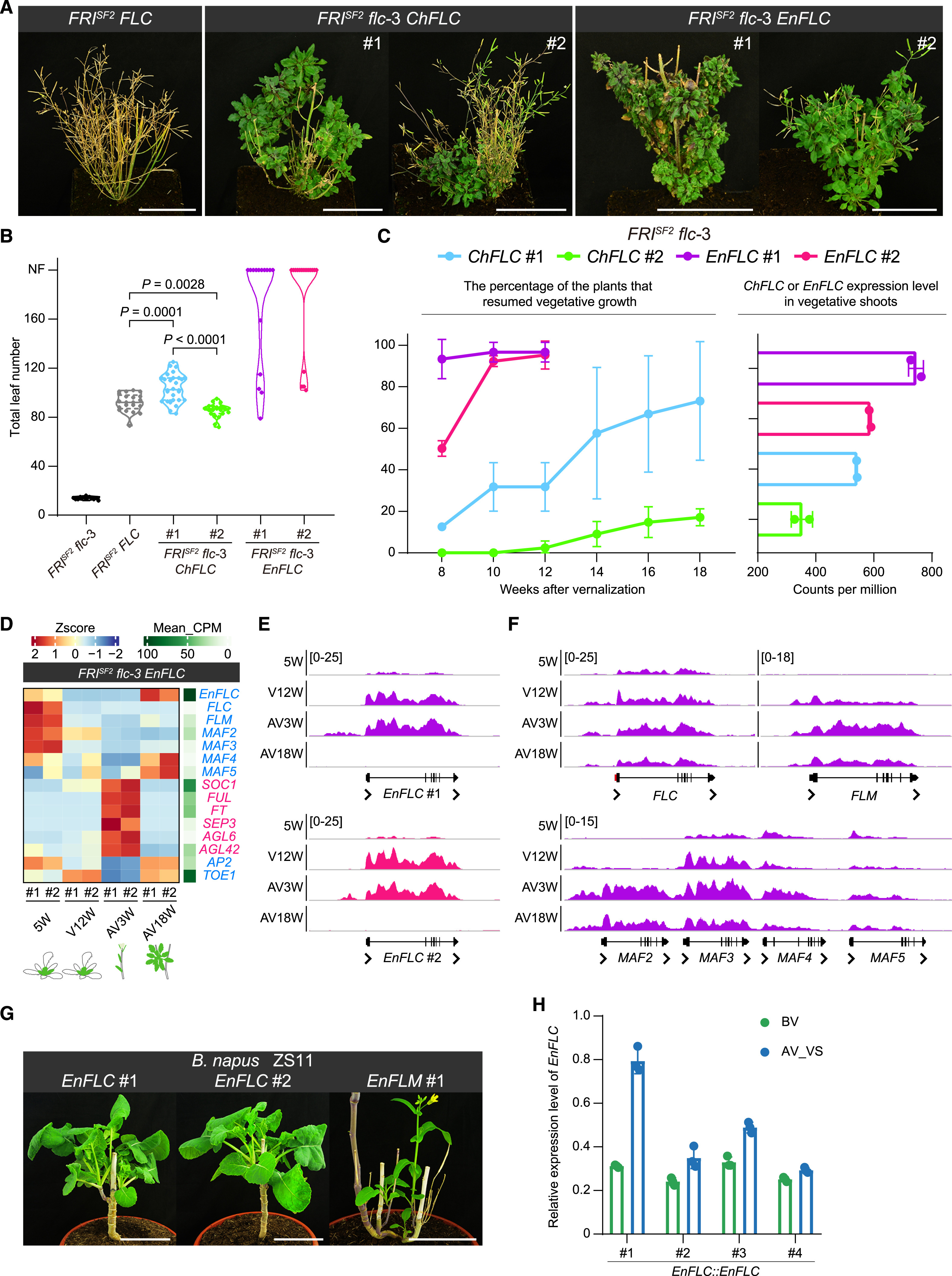
图7. 十字花科一年生植物向多果多年生开花植物的转化(图片来自原文)
这项研究揭示了植物生命周期转变的遗传机制,为培育新的多年生作物提供了理论基础。通过改变植物中类FLC基因的表达,可以培育出适应不同环境和需求的多年生作物,例如,培育多年生蔬菜可以减少农民的劳动强度,降低生产成本,并减少化肥和农药的使用,从而更加环保。培育多年生花卉可以美化环境,减少城市绿化成本,并提高城市生态效益。此外,培育多年生作物还可以提高土地利用率,减少耕作次数,并增加土壤有机质,从而提高土壤肥力。
当然,还有许多问题需要解决,如这三个基因极有可能在控制植物寿命方面也起着冗余作用,但其确切机制有待进一步研究。尽管研发人员能够在生长室条件下恢复冬生甘油菜在春化后的营养生长,但尚不清楚这些植物在田间是否能表现为真正的多果多年生植物。让我们一起期待未来研发人员的进一步研究!
翌圣助力产品
在该研究中,研究团队使用了翌圣生物PCR Mix试剂进行基因鉴定等实验:

明星产品推荐
在PCR Mix的基础上,小翌近期全新升级的快速PCR预混液(10167ES),客户体验效果好,适用于快速PCR、菌落PCR和基因型鉴定,为您的实验添砖加瓦。
|
产品定位 |
名称 |
货号 |
规格 |
|
快速PCR升级,可菌P且适用复杂模板扩增 |
2×Hieff® Ultra-Rapid II HotStart PCR Master Mix |
1 mL/5×1 mL |
客户体验反馈专区
中国科学院分子植物科学卓越创新中心
实验信息:目的片段3,000 bp
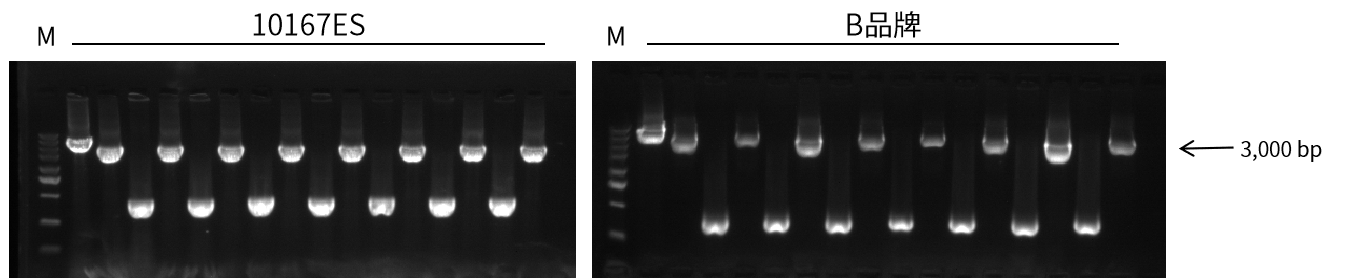
图8. 10167扩增3 kb大肠杆菌菌落,扩增效率高,产量高,性能优于竞品。延伸时间40 sec,循环数35 cycles。总反应体系10 μL。
客户评价:本产品进行大肠杆菌菌P鉴定很适合而且扩增速度很快,大大的表扬!
相关产品推荐
|
产品定位 |
名称 |
货号 |
规格 |
|
1-7片段一步法定向连接,最快5分钟完成重组反应 |
Hieff Clone® Universal II One Step Cloning Kit |
20 T/50 T |
|
|
TOPO克隆-兼容TA/平末端 |
Hieff Clone® Universal Zero TOPO TA/Blunt Cloning Kit |
20 T |
|
|
高保真PCR |
2× Hieff Canace® AdvanceFast PCR Master Mix (With Dye) |
1 mL/5×1 mL |
|
|
5 min快速完成感受态转化 |
F DH5α化学感受态 |
10×100 μL |


