
扒一扒!无血清培养添加物-常见细胞因子
无血清培养
细胞体外培养时需要在培养基中加入血清,以维持细胞的生长和存活。而血清,尤其是牛血清,成分复杂、批间差大和存在病原体污染的可能性,不确定的动物源性对细胞培养会带来显著的外源物质污染风险,因此科研人员对无血清培养基的需求日益增加,目前无血清培养基广泛应用于疫苗生产、单抗、活性蛋白等生物制品和细胞治疗等的领域中。无血清培养基中不含血清,为能够实现与含血清培养基相似的维持和促进细胞生长的功能,其中必须添加白蛋白(Albumin)、胰岛素(Insulin) 和转铁蛋白(Transferrin)等营养成分来替代血清。
生长因子在无血清培养中的作用
促细胞贴壁
许多细胞必须添加促贴壁物质才能贴壁生长,促贴壁物质一般为细胞外基质,如纤连蛋白、层粘连蛋白、玻连蛋白和胶原蛋白等。
增加细胞存活率
无血清培养基中不含有血清成分,细胞在无血清培养基中生长需要的营养成分更为单一,因此细胞的存活率可能会受到影响。添加适当的生长因子可以改善无血清培养基中的营养条件,进而提高细胞的存活率。无血清培养必需补充的细胞生长的营养物质,如胰岛素、转铁蛋白和血清白蛋白等。
促进细胞增殖和功能维持
生长因子可以促进细胞增殖,从而提高细胞的生长速度。在无血清培养基中,添加一些能够促进细胞增殖的生长因子,如表皮生长因子(EGF)、碱性成纤维细胞生长因子(bFGF)等,能够有效地促进细胞的增殖。
促进细胞分化
生长因子可以促进细胞分化,从而促进细胞的功能发挥。在无血清培养基中,添加一些能够促进细胞分化的生长因子,如转化生长因子β(TGF-β)、神经营养因子(NGF)等,能够有效地促进细胞的分化。
提高细胞表型的稳定性
无血清培养基中不含有血清成分,对于某些细胞来说,可能会导致细胞的表型不稳定。添加适当的生长因子可以提高细胞表型的稳定性,从而提高细胞的应用效果。
无血清培养基中常添加的生长因子
碱性成纤维细胞生长因子 (bFGF)
bFGF是一种肝素结合促有丝分裂蛋白,可在无血清或减血清条件下促进多种细胞的增殖。bFGF是干细胞培养中的一种关键生长因子,它不仅是重要的促有丝分裂因子,还是形态发生和分化的诱导因子。在胚胎干细胞培养中,bFGF有助于干细胞在无血清培养基中保持未分化状态。
胰岛素(Insulin)
胰岛素可维持无血清细胞培养液中细胞的生长,调节细胞对葡萄糖、氨基酸和脂肪酸的摄取、利用和贮存,同时抑制糖原、蛋白质和脂肪的分解。胰岛素也具有对抗细胞凋亡的作用。胰岛素通常以每升1-10mg的浓度添加到细胞培养基中。
表1. 胰岛素对细胞作用的分类及特点
胰岛素样生长因子-1 (IGF-1)
在无血清培养基的配制中,常用IGF-I 来替代胰岛素,IGF-I 在很低的浓度即可实现促细胞生长作用,而胰岛素需要远远高于生理浓度。IGF-1通过与IGF-1受体结合,促进蛋白质合成代谢,影响糖原代谢、DNA合成和葡萄糖摄取,具有抗凋亡作用。在无血清培养基中,IGF-1有助于细胞维持较高的活率和持续增殖的能力。
长效IGF-I(LR3 IGF-I)
Long R3 IGF-I (LR3 IGF-I) 也可称为长效IGF-I,与胰岛素相比,LR3 IGF-I添加到无血清培养基中后不容易被降解,作用更持久,因此也叫长效IGF-1。
LR3 IGF-I 是以IGF-I 为基础改造而来。改造后,LR3 IGF-I 仍具有高亲合力结合IGF-IR 的能力,因此保持着与IGF-I 相似的促细胞增殖活性。同时,改造后,LR3 IGF-I与IGF 结合蛋白( IGFBPs)的亲合力明显下降。所有哺乳动物细胞均会分泌IGFBPs,而IGFBP可结合并抑制IGF-I 的活性,而改造后的LR3 IGF-I 与IGFBPs 的亲合力比其原型分子IGF-I 下降1000 多倍,因此其活性可极大程度地避免被IGFBPs 所抑制,从而使LR3 IGF-I 的活性较IGF-I 大为提高。
LR3 IGF-1相比胰岛素,促细胞增殖能力更强,培养的细胞密度更高,细胞活力更好,且LR3 IGF-1比胰岛素在培养液中半衰期更长,生物活性更长效。
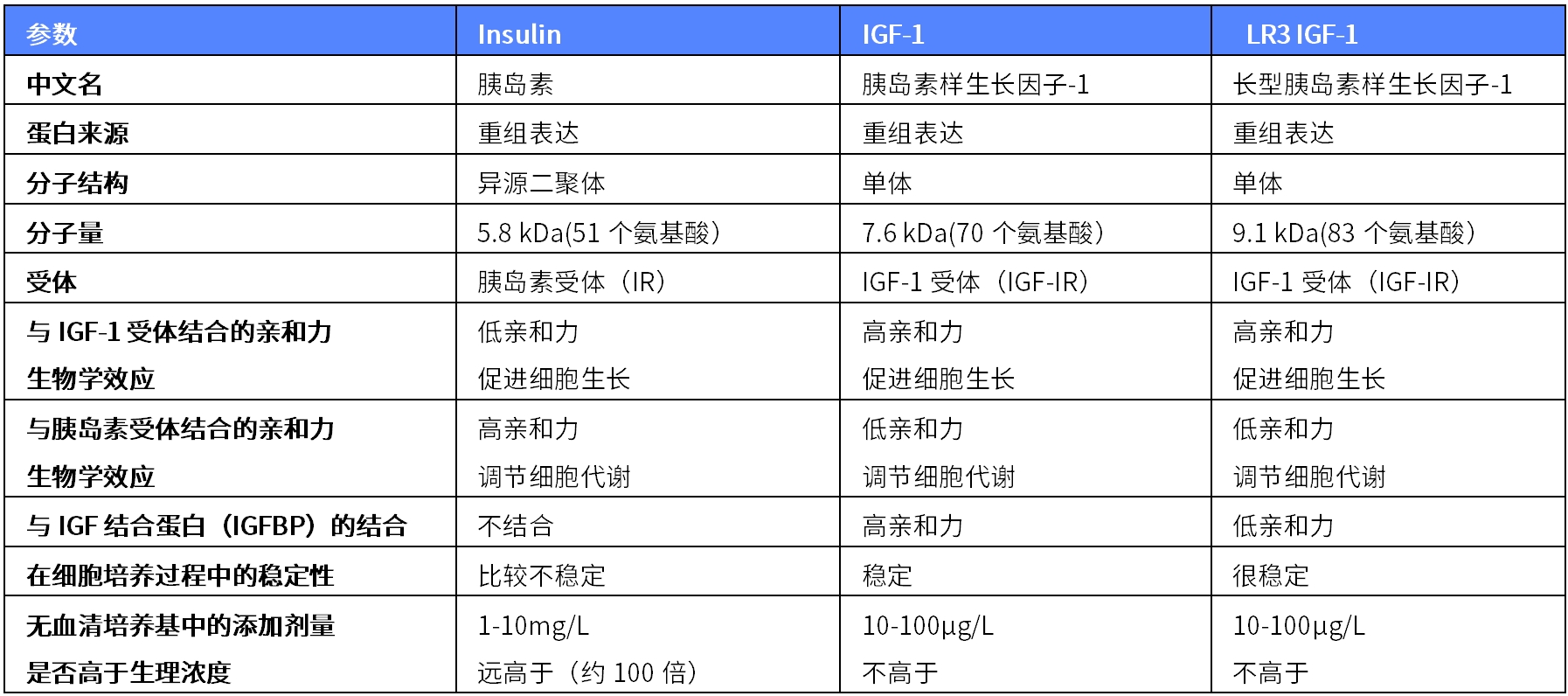
表2. 人 LR3 IGF-1、IGF-1和胰岛素的区别
|
参数 |
Insulin |
IGF-1 |
LR3 IGF-1 |
|
中文名 |
胰岛素 |
胰岛素样生长因子-1 |
长型胰岛素样生长因子-1 |
|
蛋白来源 |
重组表达 |
重组表达 |
重组表达 |
|
分子结构 |
异源二聚体 |
单体 |
单体 |
|
分子量 |
5.8 kDa(51个氨基酸) |
7.6 kDa(70个氨基酸) |
9.1 kDa(83个氨基酸) |
|
受体 |
胰岛素受体(IR) |
IGF-1受体(IGF-IR) |
IGF-1受体(IGF-IR) |
|
与IGF-1受体结合的亲和力 生物学效应 |
低亲和力 促进细胞生长 |
高亲和力 促进细胞生长 |
高亲和力 促进细胞生长 |
|
与胰岛素受体结合的亲和力 生物学效应 |
高亲和力 调节细胞代谢 |
低亲和力 调节细胞代谢 |
低亲和力 调节细胞代谢 |
|
与IGF结合蛋白(IGFBP)的结合 |
不结合 |
高亲和力 |
低亲和力 |
|
在细胞培养过程中的稳定性 |
比较不稳定 |
稳定 |
很稳定 |
|
无血清培养基中的添加剂量 是否高于生理浓度 |
1-10mg/L 远高于(约100倍) |
10-100μg/L 不高于 |
10-100μg/L 不高于 |
表皮生长因子 (EGF)
一种重要的促有丝分裂生长因子,通过与细胞表面受体结合,促进培养细胞的长期增殖。在无血清细胞或减血清条件下培养基中,诱导多种细胞的分化。
肝细胞生长因子 (HGF)
HGF是成熟实质肝细胞的一种强效、间质衍生的有丝分裂原,广泛组织和细胞类型的生长因子。HGF通过MET的跨膜酪氨酸激酶受体发出信号。HGF的活性包括诱导细胞增殖、运动、形态发生、抑制细胞生长和增强神经元存活。HGF是肝脏再生过程中至关重要的有丝分裂原。HGF能够刺激原代无血清培养的肝细胞DNA的合成,对多种组织器官的生长发育具有重要的生理调节功能。
转铁蛋白 (Transferrin)
无血清培养基为达到与含血清培养基相同或相似的细胞培养性能,其中也必须添加转铁蛋白,以实现对细胞铁离子平衡的调节,以及对细胞生长和增殖的维持。
分类:转铁蛋白以三种形态存在:脱铁(Apo)(不结合铁离子)、饱和铁(Holo)(结合两分子铁离子)和部分饱和。
如何选择:无血清培养基中多数都添加Holo-transferrin,因为Holo-transferrin 除发挥转铁作用外,自身即可作为细胞的铁源,无需在培养基中另加外源性铁。但在某些本身含铁量高的培养基(如DMEM/F12)中,则可使用Apo-transferrin 这种不含铁的转铁蛋白。
使用浓度:细胞种类不同,对转铁蛋白的需求量也不同,下表仅供参考。具体的浓度还是要根据细胞的种类、状态和想要实现的功能来通过实验确定。
表3.转铁蛋白在无血清培养基中的使用浓度
|
细胞类型 |
原代细胞 |
CHO(中国仓鼠卵巢细胞) |
杂交瘤和Vero细胞 |
|
转铁蛋白工作浓度 |
5-100mg/L |
10-50mg/mL |
5-20mg/mL |
转铁蛋白在无血清培养基中的作用:
避免细胞外环境中自由基的产生,防止细胞受到伤害
转铁蛋白可以与铁离子高亲合力结合,当被添加入无血清培养基后,在细胞外的培养环境中基本不会存在游离的铁离子,也就不会发生自由基反应,故可避免自由基对细胞的损伤。
维持细胞的生长和增殖
细胞内有很多重要的蛋白,如血红素、核糖核苷酸还原酶、DNA解旋酶、DNA聚合酶、DNA引物酶等,需要结合铁离子后才能发挥其活性,这些蛋白基本上都与DNA 的复制、修复以及DNA 的稳定性有关,也就是说它们影响着细胞的生长和增殖。转铁蛋白通过与其在细胞表面的受体结合,将铁离子转运并释放到细胞内后,这些铁离子会与需铁蛋白结合,然后发挥后者维持细胞生长和增殖的功能。如果无血清培养基中不含转铁蛋白,细胞内的需铁蛋白将无法获得铁离子,会失去活性,导致细胞生长停滞或死亡。
维持细胞内的铁平衡
在细胞培养中,并不是铁越多越好,而应当是在适当的时间获得适量的铁。如果细胞内的铁过多,也跟细胞外环境一样,铁会诱导产生破坏力极强的羟基自由基。而如果细胞内铁过少,则不足以维持细胞的生长和增殖。转铁蛋白调节细胞内的铁平衡,与细胞表面的转铁蛋白受体有关。细胞表面转铁蛋白受体的数目不是固定不变的,而是随着细胞对铁的需求程度而上调或下调,因而可自主影响细胞对铁的摄取量一般而言,静止未活化的细胞表面转铁蛋白受体相对较少,而活化增殖的细胞表面转铁蛋白受体数目显著增加。此外,在细胞培养中,转铁蛋白受体数目与培养环境中铁的含量也密切相关。培养环境中的含铁介质比较多,则细胞表面转铁蛋白受体的数目会减少,从而避免细胞摄入过量的铁。
白蛋白(HSA)
白蛋白是许多无血清细胞培养的重要组成部分,例如杂交瘤或中国仓鼠卵巢(CHO)细胞的系统。白蛋白具有多种生理功能,包括:金属离子结合剂、自由基清除剂、渗透压和pH值的调节剂、生长因子的运载体、表面活性剂以及营养剂等。
纤维连接蛋白(fibronectin,FN)
纤维连接蛋白添加到培养基中,替换或降低血清使用量,促进细胞贴壁、迁移、移动生长、维持细胞状态。具体作用如下:
纤维连接蛋白可以与细胞表面整合素等受体结合,促进细胞与培养皿的粘附,这是细胞成功培养的前提。特别是一些黏附力较弱的细胞,添加纤维连接蛋白可以显著提高其培养成功率。
纤维连接蛋白除了增强细胞附着外,还可以通过与细胞表面受体的相互作用,激活下游信号转导通路,促进细胞周期进程,实现细胞的增殖。
维持细胞形态。纤维连接蛋白与细胞结合后,可以影响细胞骨架的排列和细胞形态的维持。此作用对于某些依赖于reshape的细胞功能和分化很重要。
促进细胞迁移。纤维连接蛋白在体内发挥着引导细胞迁移的作用。在体外培养中可以形成化学梯度以诱导细胞的定向迁移,这对研究细胞转移机制很重要。
诱导细胞分化。不同浓度和构象的纤维连接蛋白,可以与不同类型的细胞相互作用,展示不同的生物学效应,诱导细胞向特定的方向分化。这在定向诱导多潜能细胞分化中很重要。
相关产品推荐
|
产品名称 |
货号 |
规格 |
|
91330ES |
10μg/100μg |
|
|
40112ES |
25mg/100mg |
|
|
92211ES |
10μg/100μg |
|
|
92210ES |
5μg/100μg |
|
|
92708ES |
100μg/500μg |
|
|
92055ES |
10μg/50μg |
|
|
92288ES |
100 mg/500 mg |
|
|
92289ES |
100 mg/500 mg |
|
|
40133ES |
100 mg/500 mg |
|
|
92618ES |
1g/5g |
|
|
20901ES |
1g/5g |
|
|
40113ES |
1mg/10mg |

