
上新|热稳定、低宿主残留的RNase HII来咯!
如果您想搜的是RNase H,可点击链接获取RNase H产品详情
RNase HII是什么酶
RNase HII是一种核糖核酸内切酶,识别并切割嵌入DNA双链中的单个核糖核昔酸5端位点的磷酸二醋键,切割后产生一个 5′ 端磷酸基团和一个 3′ 端羟基(图1)。但RNase HII对单链RNA切割活性极低,对dsDNA、ssDNA无切割活性。RNase HII在50°C~75°C间具有活性,在70~75°C具有最佳活性。由于RNase HII具有在DNA掺入单个核糖核苷酸位点处进行单点切割而对dsDNA、ssDNA无切割活性的特性,RNase HII常用于启动反应(引物激活并延伸)的“开关”,从而实现特异性扩增,在RNase HII依赖性PCR(rhPCR)、环介导等温扩增技术(LAMP)、冈崎片段(Okazaki fragment)部分RNA降解等应用广泛。
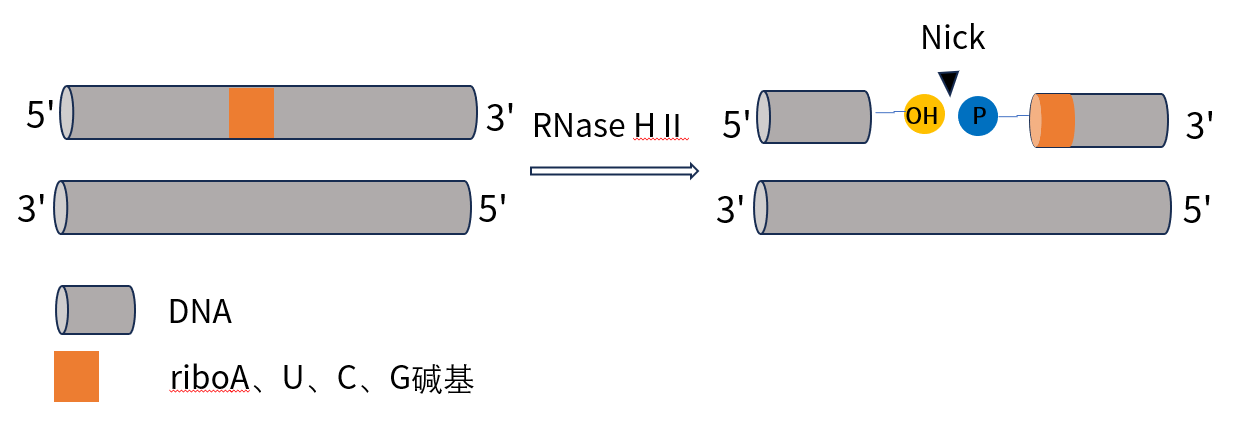
图1. RNase HII反应原理示意图
RNase HII的功能应用
依赖于Rnase HII的PCR (rhPCR)
rhPCR即依赖于RNase HII的PCR,在PCR的基础上增加了RNase HII和封闭式可切割rhPCR引物。
rhPCR利用了RNase HII的特殊性质,即可以特异性地水解DNA-RNA杂合链中的RNA,但不能水解单链或双链DNA或RNA中的磷酸二酯键,不能消化单链或双链DNA或RNA。因此只有当互补靶DNA碱基存在时,RNase HII才能消化DNA-RNA杂交链从而允许引物延伸。由于仅在与互补靶序列紧密结合时才发生rhPCR引物的裂解切割,故极大地提高了反应的准确性。RNase HII是唯一能够通过水解核糖核酸5' 端的磷酸二酯键来启动无突变去除核糖核苷酸过程的酶。rhPCR因其特异性、灵敏度和重复性,在单核苷酸多态性(SNP)、基因分型、多靶标同时检测、环境核酸检测等方面具有一定的优势。(戳连接,了解详情)
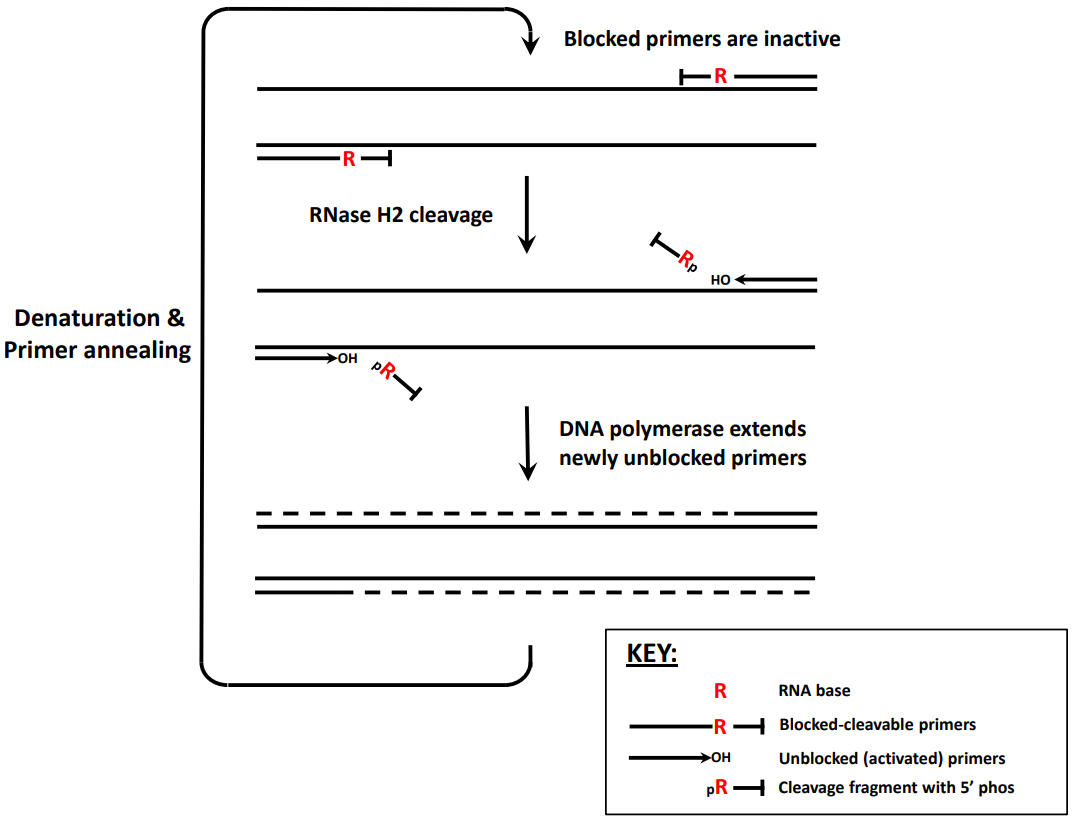
图2. rhPCR原理图[1]
环介导等温扩增技术(LAMP)
环介导等温扩增技术(LAMP)是一种简便、快速的基因扩增方法,能在等温条件下,短时间内进行核酸扩增。但由于在常温配制过程中,DNA聚合酶已经开始工作,形成非特异性的错配,容易产生少量错配和引物二聚体,这些轻微的污染都会造成假阳性。
将RNase HII应用于LAMP扩增体系后,能有效解决LAMP技术的假阳性问题,为LAMP技术更加广泛的应用于临床诊断。如图3所示,利用Rnase HII引物激活的LAMP方法(PA-LAMP),当引物与靶ssDNA配对时,Rnase HII酶特异性切割引物上的RNA,激活引物,从而允许引物延伸,实现特异性扩增,有效减少体系中的假阳性。
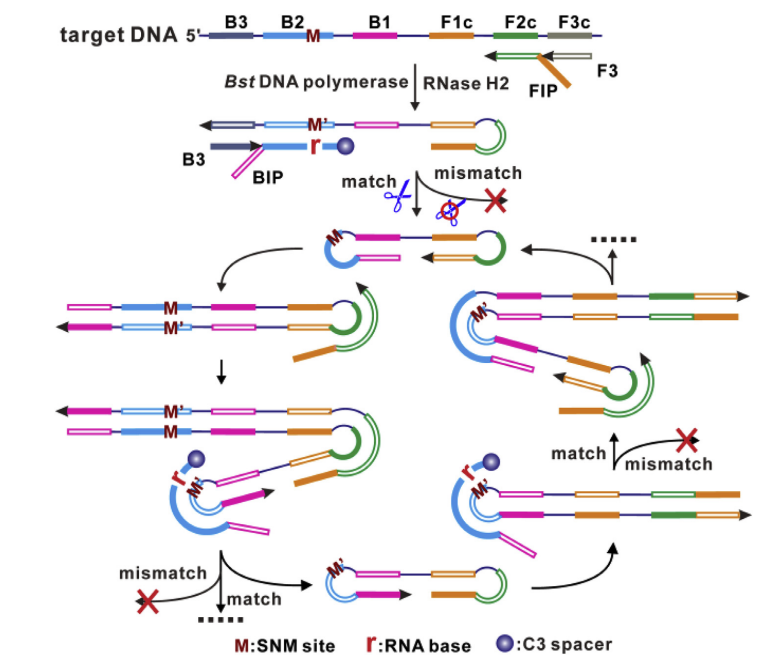
图3. PA-LAMP原理示意图[2]
翌圣RNase HⅡ
翌圣的RNase HⅡ(Cat#14539)来源于深渊热球菌 (Pyrococcus abyssi, P.a.),无核酸外切酶、切口酶、RNase残留,低宿主残留,有效减少体系假阳性。翌圣RNase HⅡ极度耐热,95°C反应30 min后,仍保持活性,可与rhPCR各反应体系兼容,也适用于LAMP等。
E. coli基因组DNA残留< 0.5copies/100 U
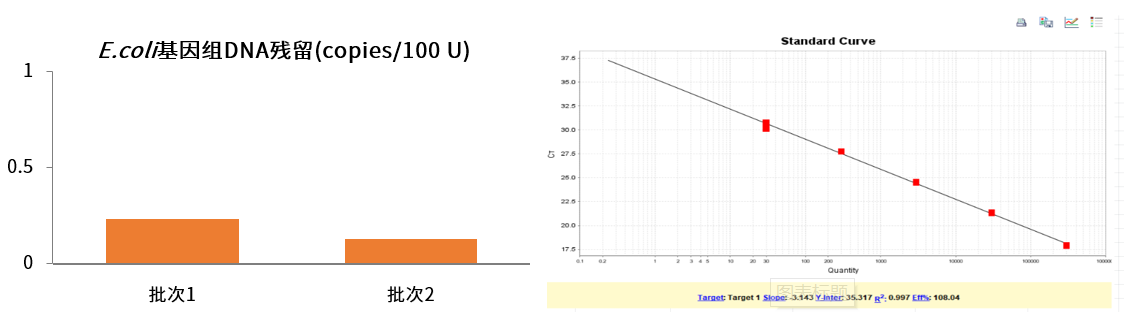
图4. E. coli基因组DNA残留检测
95℃耐热性测试
对RNase HⅡ进行95℃加热0~45 min后,测试RNase HⅡ的酶活结果,结果表明翌圣RNase HⅡ加热45 min后,酶活几乎没有损失。
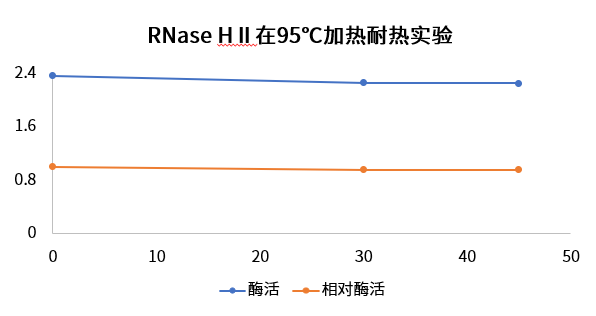
图5. 95℃耐热测试
无核酸外切酶、切口酶、RNase残留(1 U投入量)
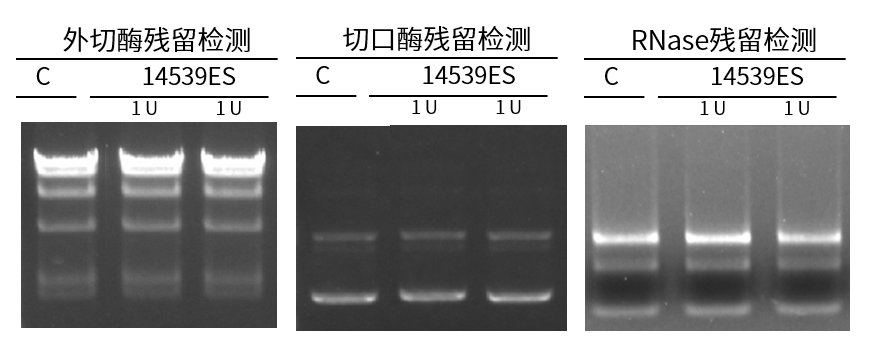
图6. 核酸外切酶、切口酶、RNase残留检测结果
相关产品推荐
|
产品应用 |
产品名称 |
产品货号 |
|
rhPCR、LAMP |
RNase HII(2 U/μL) |
|
|
rhPCR |
Hieff® Taq DNA Polymerase DNA聚合酶 |
|
|
RT-LAMP |
Hieff® Bst Plus DNA Polymerase (40 U/μL) |
|
|
Hifair® Ⅲ Reverse Transcriptase 第三代耐热逆转录酶 |
参考文献
[1] Dobosy JR, Rose SD, Beltz KR, Rupp SM, Powers KM, Behlke MA, Walder JA. RNase H-dependent PCR (rhPCR): improved specificity and single nucleotide polymorphism detection using blocked cleavable primers. BMC Biotechnol. 2011 Aug 10;11:80. doi: 10.1186/1472-6750-11-80.
[2] Du W F , Ge J H , Li J J ,et al.Single-step, high-specificity detection of single nucleotide mutation by primer-activatable loop-mediated isothermal amplification (PA-LAMP)[J].Analytica chimica acta, 2019(1050-):1050.DOI:10.1016/j.aca.2018.10.068.

